Das könnte Ihnen auch gefallen
- Seasonally Dry Tropical Forests: Ecology and ConservationVon EverandSeasonally Dry Tropical Forests: Ecology and ConservationNoch keine Bewertungen
- Acta Protozoologica 49 1 3Dokument12 SeitenActa Protozoologica 49 1 3Fernanda Charqueño CelisNoch keine Bewertungen
- 81845918Dokument13 Seiten81845918jime joel ureta porrasNoch keine Bewertungen
- WernerEtAl 2011 IsTheResilienceOfEpiphyteAssemblagesDokument6 SeitenWernerEtAl 2011 IsTheResilienceOfEpiphyteAssemblagesLau Rodríguez LavínNoch keine Bewertungen
- tmp69 TMPDokument12 Seitentmp69 TMPFrontiersNoch keine Bewertungen
- 1996 Jackson EtalDokument24 Seiten1996 Jackson Etalpashkito2014Noch keine Bewertungen
- Uezu, Beyer and Metzger 2008Dokument16 SeitenUezu, Beyer and Metzger 2008Vitoria Duarte DerissoNoch keine Bewertungen
- Ekoungoulou Et Al., 2017Dokument13 SeitenEkoungoulou Et Al., 2017Romeo EkoungoulouNoch keine Bewertungen
- Fauna DarlynDokument14 SeitenFauna DarlynDARLYN ASNIT LOPEZ JIMENEZNoch keine Bewertungen
- Forest Ecology and ManagementDokument7 SeitenForest Ecology and ManagementClaudio SantanaNoch keine Bewertungen
- Cairns 1997 Root Biomass Allocation in The Worlds Upland ForestsDokument12 SeitenCairns 1997 Root Biomass Allocation in The Worlds Upland Forestsfatmi noor'anNoch keine Bewertungen
- Morphofunctional Groups BryophytesDokument11 SeitenMorphofunctional Groups BryophytesLuciana Silva da CostaNoch keine Bewertungen
- Lohmus 2011 Silviculture As A Disturbance Regime The Effects of Clear Cutting Planting and Thinning On Polypore Communities in Mixed ForestsDokument10 SeitenLohmus 2011 Silviculture As A Disturbance Regime The Effects of Clear Cutting Planting and Thinning On Polypore Communities in Mixed Forestsvonjavier80Noch keine Bewertungen
- Understanding Mixed Woody-Herbaceous EcosystemsDokument16 SeitenUnderstanding Mixed Woody-Herbaceous Ecosystemssandrino1091Noch keine Bewertungen
- 07 Schnitzler Etal 2007 BiolconservDokument11 Seiten07 Schnitzler Etal 2007 BiolconservprabodhdrdoNoch keine Bewertungen
- Edge-Interior Differences in Vegetation Structure and Composition in A Chaco Semi-Arid Forest, ArgentinaDokument9 SeitenEdge-Interior Differences in Vegetation Structure and Composition in A Chaco Semi-Arid Forest, ArgentinaAnonymous ZON7qMNoch keine Bewertungen
- Species Diversity of Canopy Versus Understory Trees in A Neotropical Forest: Implications For Forest Structure, Function and MonitoringDokument13 SeitenSpecies Diversity of Canopy Versus Understory Trees in A Neotropical Forest: Implications For Forest Structure, Function and MonitoringLight WhiteNoch keine Bewertungen
- Erdmann2012 Article RegionalFactorsRatherThanForesDokument13 SeitenErdmann2012 Article RegionalFactorsRatherThanForesSebastian MartinezNoch keine Bewertungen
- Paper 08Dokument8 SeitenPaper 08vitaNoch keine Bewertungen
- Costa Et Al. 2008 Gradients Within Gradients Central Amazonian ForestDokument11 SeitenCosta Et Al. 2008 Gradients Within Gradients Central Amazonian ForestccruzvasconcelosNoch keine Bewertungen
- 2011, Matthias Schleuning Et. Al, La Dinámica Del Río Da Forma A La Diversidad Clonal y La GenéticaDokument14 Seiten2011, Matthias Schleuning Et. Al, La Dinámica Del Río Da Forma A La Diversidad Clonal y La GenéticaVicky Huamán QquellónNoch keine Bewertungen
- Plant Functional Groups Within A Tropical Forest Exhibit Different Wood Functional AnatomyDokument10 SeitenPlant Functional Groups Within A Tropical Forest Exhibit Different Wood Functional AnatomyAriadne Cristina De AntonioNoch keine Bewertungen
- Environmental Conditions Affecting Spiders in GrasDokument11 SeitenEnvironmental Conditions Affecting Spiders in GrasRaymond OrnopiaNoch keine Bewertungen
- Carbon Stock and Woody Species Diversity in HomegardenDokument12 SeitenCarbon Stock and Woody Species Diversity in HomegardenNuring SangkelatNoch keine Bewertungen
- PDF 127342 60737Dokument11 SeitenPDF 127342 60737raja sohaib qayyumNoch keine Bewertungen
- Fine-Scale - Topography ALICEDokument10 SeitenFine-Scale - Topography ALICEluz marina mulato campoNoch keine Bewertungen
- Global analysis of root distributions across terrestrial biomesDokument23 SeitenGlobal analysis of root distributions across terrestrial biomesBrandon Vander DeckenNoch keine Bewertungen
- Brown Gillespieetal 1989Dokument11 SeitenBrown Gillespieetal 1989Muhammad AhmadNoch keine Bewertungen
- Regulation of Leaf Life-Span and Nutrient-Use Efficiency of Metrosideros Polymorpha Trees at Two Extremes of A Long Chronosequence in HawaiiDokument9 SeitenRegulation of Leaf Life-Span and Nutrient-Use Efficiency of Metrosideros Polymorpha Trees at Two Extremes of A Long Chronosequence in HawaiiDeliane OliveiraNoch keine Bewertungen
- Adjalla Et Al 2021, Soil Seed Bank Characteristics Along A Gradient of Past Human DisturbancesDokument13 SeitenAdjalla Et Al 2021, Soil Seed Bank Characteristics Along A Gradient of Past Human DisturbancesADJALLANoch keine Bewertungen
- (Q1) Carpenter2012 Article BiodiversityOfSoilMacrofaunaInDokument26 Seiten(Q1) Carpenter2012 Article BiodiversityOfSoilMacrofaunaInSalma FahimaNoch keine Bewertungen
- Biodiversity de Jong Et Al, Effect of Forest ManagementDokument12 SeitenBiodiversity de Jong Et Al, Effect of Forest ManagementMán EdeNoch keine Bewertungen
- Chave Et Al-2005 Ecuaciones AlometricasDokument13 SeitenChave Et Al-2005 Ecuaciones AlometricasOswaldo JadánNoch keine Bewertungen
- Apaza-Quevedo Et Al 2015 (Edge Effects)Dokument10 SeitenApaza-Quevedo Et Al 2015 (Edge Effects)BeatrizNoch keine Bewertungen
- Buckton2019 - Leaf and WDDokument12 SeitenBuckton2019 - Leaf and WDBeatrizNoch keine Bewertungen
- Agriculture, Ecosystems and Environment: A A B C CDokument11 SeitenAgriculture, Ecosystems and Environment: A A B C CPutriNoch keine Bewertungen
- Kessler Et Al 2014. Elevational Petterns of PolylepisDokument12 SeitenKessler Et Al 2014. Elevational Petterns of PolylepisOsir Daygor VidalNoch keine Bewertungen
- CarbonDokument8 SeitenCarbonobsa tayeNoch keine Bewertungen
- Art 13. Impactos de Los Bosques y La Forestación Sobre Los Servicios Hidrológicos en Los Andes Una Revisión SistemáticaDokument16 SeitenArt 13. Impactos de Los Bosques y La Forestación Sobre Los Servicios Hidrológicos en Los Andes Una Revisión Sistemáticamonica oviedoNoch keine Bewertungen
- tmpB5B9 TMPDokument12 SeitentmpB5B9 TMPFrontiersNoch keine Bewertungen
- Forest Ecology and Management: Yik-Hei Sung, Nancy E. Karraker, Billy C.H. HauDokument7 SeitenForest Ecology and Management: Yik-Hei Sung, Nancy E. Karraker, Billy C.H. HauAlejandra CardonaNoch keine Bewertungen
- 2.ecological Trends in The Wood Anatomy of Vaccinioideae (Ericaceae S.L.)Dokument11 Seiten2.ecological Trends in The Wood Anatomy of Vaccinioideae (Ericaceae S.L.)Eileen Caroline Ruiz NunezNoch keine Bewertungen
- Coronado Molina2004Dokument8 SeitenCoronado Molina2004fargotty210026Noch keine Bewertungen
- Effect of Climate and Physiography On The Density of Tree and Shrub Species in Northwest MexicoDokument13 SeitenEffect of Climate and Physiography On The Density of Tree and Shrub Species in Northwest MexicoPablo M. AntúnezNoch keine Bewertungen
- Biological Conservation: Renata Durães, Luis Carrasco, Thomas B. Smith, Jordan KarubianDokument9 SeitenBiological Conservation: Renata Durães, Luis Carrasco, Thomas B. Smith, Jordan KarubianSelsa ArtikaNoch keine Bewertungen
- Moss Diversity Along Altitudinal Gradients in Mt. MakilingDokument3 SeitenMoss Diversity Along Altitudinal Gradients in Mt. MakilingRamil AlcalaNoch keine Bewertungen
- Tropical Forests Are Not Flat: How Mountains Affect Herbivore DiversityDokument10 SeitenTropical Forests Are Not Flat: How Mountains Affect Herbivore DiversityMarina Trillo GabaldonNoch keine Bewertungen
- 17 Kerns 2003 Phyto Concentration Soil Properties Vegetation PinusDokument11 Seiten17 Kerns 2003 Phyto Concentration Soil Properties Vegetation PinusMarcel FumiyaNoch keine Bewertungen
- Luize Etal 2015 Inundacao e Variedade VegetacaoDokument6 SeitenLuize Etal 2015 Inundacao e Variedade Vegetacaogomes.pvoliveiraNoch keine Bewertungen
- The Legacy of Biogeographic History On The Composition and Structure of Andean ForestsDokument11 SeitenThe Legacy of Biogeographic History On The Composition and Structure of Andean ForestsDiego Andres Moreno GaonaNoch keine Bewertungen
- Determinants and Correlates of Above Ground Biomass in A Secondary Hillside Rainforest in Central VietnamDokument27 SeitenDeterminants and Correlates of Above Ground Biomass in A Secondary Hillside Rainforest in Central VietnamTuyen TruongNoch keine Bewertungen
- Predictors of Forest Fragmentation Sensitivity in Neotropical Vertebrates - A Quantitative ReviewDokument8 SeitenPredictors of Forest Fragmentation Sensitivity in Neotropical Vertebrates - A Quantitative ReviewsamuelvelasquezfajardoNoch keine Bewertungen
- Brooks2008 Temperatura Del Suelo Del Bosque y Humedad Relativa Luego de La Extracción de Madera en El Sur de Nueva InglaterraDokument9 SeitenBrooks2008 Temperatura Del Suelo Del Bosque y Humedad Relativa Luego de La Extracción de Madera en El Sur de Nueva Inglaterramiguel angelNoch keine Bewertungen
- Genetic Adaptation vs. Ecophysiological Plasticity of Photosynthetic-Related Traits in Young Picea Glauca Trees Along A Regional Climatic GradientDokument15 SeitenGenetic Adaptation vs. Ecophysiological Plasticity of Photosynthetic-Related Traits in Young Picea Glauca Trees Along A Regional Climatic GradientJhon Nieto VargasNoch keine Bewertungen
- Journal of Animal Ecology - 2009 - Keller - The Importance of Environmental Heterogeneity For Species Diversity andDokument10 SeitenJournal of Animal Ecology - 2009 - Keller - The Importance of Environmental Heterogeneity For Species Diversity andsamanthavieira.s39Noch keine Bewertungen
- ThesisDokument14 SeitenThesischerry m cabatbatNoch keine Bewertungen
- Bezand Homeier 2017 GCBDokument22 SeitenBezand Homeier 2017 GCBCarla MaldonadoNoch keine Bewertungen
- Brehm, Et Al. 2006Dokument15 SeitenBrehm, Et Al. 2006zulmakqa4819Noch keine Bewertungen
- Austral Ecology - 2008 - Lusk - Survival and Growth of Seedlings of 12 Chilean Rainforest Trees in Two Light EnvironmentsDokument10 SeitenAustral Ecology - 2008 - Lusk - Survival and Growth of Seedlings of 12 Chilean Rainforest Trees in Two Light EnvironmentsAlén MirandaNoch keine Bewertungen
- Bats of The Chilean TemperateDokument24 SeitenBats of The Chilean TemperateMassiel Aravena GavilanNoch keine Bewertungen
- Komyiama Et Al. (2008) - Allometry, Biomass, and Productivity of Mangrove Forests - A Review PDFDokument10 SeitenKomyiama Et Al. (2008) - Allometry, Biomass, and Productivity of Mangrove Forests - A Review PDFcarlitos254Noch keine Bewertungen
- BioCarbon Tracker Report - PNSLDokument9 SeitenBioCarbon Tracker Report - PNSLcarlitos254Noch keine Bewertungen
- Green Carbon GuidebookDokument14 SeitenGreen Carbon Guidebookcarlitos254Noch keine Bewertungen
- Geonoma SeleriDokument2 SeitenGeonoma Selericarlitos254Noch keine Bewertungen
- Cifuentes Jara Et Al 2013 Inventory of Volume and Biomass Tree Allometric Equations-LibreDokument80 SeitenCifuentes Jara Et Al 2013 Inventory of Volume and Biomass Tree Allometric Equations-Librecarlitos254Noch keine Bewertungen
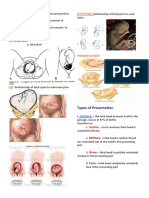
- The 4Ps of Labor: Passenger, Passageway, Powers, and PlacentaDokument4 SeitenThe 4Ps of Labor: Passenger, Passageway, Powers, and PlacentaMENDIETA, JACQUELINE V.Noch keine Bewertungen
- Ethanox 330 Antioxidant: Safety Data SheetDokument7 SeitenEthanox 330 Antioxidant: Safety Data Sheetafryan azhar kurniawanNoch keine Bewertungen
- Midnight HotelDokument7 SeitenMidnight HotelAkpevweOghene PeaceNoch keine Bewertungen
- The Ramayana and The Sacred Palm Trees of Sumeria, Mesopotamia, Assyria and PhoeniciaDokument7 SeitenThe Ramayana and The Sacred Palm Trees of Sumeria, Mesopotamia, Assyria and PhoeniciaNeeta RainaNoch keine Bewertungen
- DC Rebirth Reading Order 20180704Dokument43 SeitenDC Rebirth Reading Order 20180704Michael MillerNoch keine Bewertungen
- Good Paper On Time SerisDokument15 SeitenGood Paper On Time SerisNamdev UpadhyayNoch keine Bewertungen
- Journal of Electronic MaterialsDokument10 SeitenJournal of Electronic MaterialsSanjib BaglariNoch keine Bewertungen
- Postmodern Dystopian Fiction: An Analysis of Bradbury's Fahrenheit 451' Maria AnwarDokument4 SeitenPostmodern Dystopian Fiction: An Analysis of Bradbury's Fahrenheit 451' Maria AnwarAbdennour MaafaNoch keine Bewertungen
- Salzer Panel Accessories Price List - 01st January 2019Dokument40 SeitenSalzer Panel Accessories Price List - 01st January 2019Chandra SekaranNoch keine Bewertungen
- The Principle of Subsidiarity and Catholic Ecclesiology - ImplicatDokument218 SeitenThe Principle of Subsidiarity and Catholic Ecclesiology - ImplicatJonathanKiehlNoch keine Bewertungen
- Public Policy For Nigerian EnviromentDokument11 SeitenPublic Policy For Nigerian EnviromentAbishai Auta GaiyaNoch keine Bewertungen
- MCQ CH 5-Electricity and Magnetism SL Level: (30 Marks)Dokument11 SeitenMCQ CH 5-Electricity and Magnetism SL Level: (30 Marks)Hiya ShahNoch keine Bewertungen
- Causes of WWI: Alliances, Militarism & AssassinationDokument4 SeitenCauses of WWI: Alliances, Militarism & AssassinationJoshua De Leon TuasonNoch keine Bewertungen
- MAT 1100 Mathematical Literacy For College StudentsDokument4 SeitenMAT 1100 Mathematical Literacy For College StudentsCornerstoneFYENoch keine Bewertungen
- Magnetic Suspension System With Electricity Generation Ijariie5381Dokument11 SeitenMagnetic Suspension System With Electricity Generation Ijariie5381Jahnavi ChinnuNoch keine Bewertungen
- Gender and DelinquencyDokument26 SeitenGender and DelinquencyCompis Jenny-annNoch keine Bewertungen
- Diagram Illustrating The Globalization Concept and ProcessDokument1 SeiteDiagram Illustrating The Globalization Concept and ProcessAnonymous hWHYwX6Noch keine Bewertungen
- INDIA'S DEFENCE FORCESDokument3 SeitenINDIA'S DEFENCE FORCESJanardhan ChakliNoch keine Bewertungen
- Entrepreneurship - Quarter 2 - Week 1-3 - 4 M's of Production and - Business ModelDokument6 SeitenEntrepreneurship - Quarter 2 - Week 1-3 - 4 M's of Production and - Business ModelJude Del RosarioNoch keine Bewertungen
- Bracketing MethodsDokument13 SeitenBracketing Methodsasd dsa100% (1)
- Mini Lecture and Activity Sheets in English For Academic and Professional Purposes Quarter 4, Week 5Dokument11 SeitenMini Lecture and Activity Sheets in English For Academic and Professional Purposes Quarter 4, Week 5EllaNoch keine Bewertungen
- Dukic WarehouselayoutsDokument14 SeitenDukic Warehouselayoutsrohitkamath7Noch keine Bewertungen
- SalutogenicshandbookDokument16 SeitenSalutogenicshandbookAna EclipseNoch keine Bewertungen
- HOTC 1 TheFoundingoftheChurchandtheEarlyChristians PPPDokument42 SeitenHOTC 1 TheFoundingoftheChurchandtheEarlyChristians PPPSuma HashmiNoch keine Bewertungen
- Present Tense Exercises. Polish A1Dokument6 SeitenPresent Tense Exercises. Polish A1Pilar Moreno DíezNoch keine Bewertungen
- Prospectus2023 24 PDFDokument332 SeitenProspectus2023 24 PDFramgharia sameerNoch keine Bewertungen
- Group Handling - Pre Registration Activity: Submited To-Submitted byDokument12 SeitenGroup Handling - Pre Registration Activity: Submited To-Submitted byharshal kushwahNoch keine Bewertungen
- Research Paper On The Hells AngelsDokument6 SeitenResearch Paper On The Hells Angelsfvg2xg5r100% (1)
- M04 - Conditional Obligations and Reciprocal Obligations PDFDokument12 SeitenM04 - Conditional Obligations and Reciprocal Obligations PDFTam GarciaNoch keine Bewertungen
- Compilation of Activities UBCVDokument13 SeitenCompilation of Activities UBCVRenell Vincent RamosNoch keine Bewertungen
- Why We Die: The New Science of Aging and the Quest for ImmortalityVon EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityBewertung: 3.5 von 5 Sternen3.5/5 (2)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisVon EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisBewertung: 3.5 von 5 Sternen3.5/5 (2)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceVon EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceBewertung: 4.5 von 5 Sternen4.5/5 (515)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsVon EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsBewertung: 4.5 von 5 Sternen4.5/5 (4)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyVon EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyBewertung: 3.5 von 5 Sternen3.5/5 (31)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorVon EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNoch keine Bewertungen
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindVon EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindBewertung: 4.5 von 5 Sternen4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondVon EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondBewertung: 4 von 5 Sternen4/5 (3)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeVon EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNoch keine Bewertungen
- The Lives of Bees: The Untold Story of the Honey Bee in the WildVon EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildBewertung: 4.5 von 5 Sternen4.5/5 (44)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessVon Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessBewertung: 4 von 5 Sternen4/5 (33)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldVon EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldBewertung: 4.5 von 5 Sternen4.5/5 (18)
- Why We Sleep: Unlocking the Power of Sleep and DreamsVon EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsBewertung: 4.5 von 5 Sternen4.5/5 (2083)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceVon EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNoch keine Bewertungen
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesVon EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesBewertung: 4.5 von 5 Sternen4.5/5 (396)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedVon EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedBewertung: 4 von 5 Sternen4/5 (11)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceVon EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceBewertung: 4 von 5 Sternen4/5 (632)
- Superlative: The Biology of ExtremesVon EverandSuperlative: The Biology of ExtremesBewertung: 4.5 von 5 Sternen4.5/5 (51)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesVon EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesBewertung: 3.5 von 5 Sternen3.5/5 (56)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeVon EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeBewertung: 4 von 5 Sternen4/5 (523)
- Summary of Robert M. Sapolsky's DeterminedVon EverandSummary of Robert M. Sapolsky's DeterminedBewertung: 5 von 5 Sternen5/5 (2)
- Lymph & Longevity: The Untapped Secret to HealthVon EverandLymph & Longevity: The Untapped Secret to HealthBewertung: 4.5 von 5 Sternen4.5/5 (13)
- Inside of a Dog: What Dogs See, Smell, and KnowVon EverandInside of a Dog: What Dogs See, Smell, and KnowBewertung: 4 von 5 Sternen4/5 (390)
- Fearfully and Wonderfully: The Marvel of Bearing God's ImageVon EverandFearfully and Wonderfully: The Marvel of Bearing God's ImageBewertung: 5 von 5 Sternen5/5 (40)
- Mitochondria and the Future of Medicine: The Key to Understanding Disease, Chronic Illness, Aging, and Life ItselfVon EverandMitochondria and the Future of Medicine: The Key to Understanding Disease, Chronic Illness, Aging, and Life ItselfBewertung: 4.5 von 5 Sternen4.5/5 (98)
- Gathering Moss: A Natural and Cultural History of MossesVon EverandGathering Moss: A Natural and Cultural History of MossesBewertung: 4.5 von 5 Sternen4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsVon EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNoch keine Bewertungen
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationVon EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationBewertung: 4 von 5 Sternen4/5 (1)