Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Ravi Gupta, S. R. Pandi-Perumal, Ahmed S. BaHammam - Clinical Atlas of Polysomnography (2018, Apple Academic Press)Dokument547 SeitenRavi Gupta, S. R. Pandi-Perumal, Ahmed S. BaHammam - Clinical Atlas of Polysomnography (2018, Apple Academic Press)psicosmosNoch keine Bewertungen
- Murder MysteryDokument12 SeitenMurder Mysteryapi-272999528Noch keine Bewertungen
- Duchenne Muscular DystrophyDokument32 SeitenDuchenne Muscular DystrophyMr.P.Ramesh, Faculty of Physiotherapy, SRUNoch keine Bewertungen
- Flavone Synthesis ThesisDokument300 SeitenFlavone Synthesis ThesisJamal Rafique100% (1)
- Experimental and Quasi-Experimental Designs For Generalized Causal Inference PDFDokument81 SeitenExperimental and Quasi-Experimental Designs For Generalized Causal Inference PDFGonçalo SampaioNoch keine Bewertungen
- Semi-Detailed Lesson Plan For Cellular RespirationDokument2 SeitenSemi-Detailed Lesson Plan For Cellular RespirationZaifel Pacillos100% (5)
- 2007 Sleep in Children With Autistic Spectrum Disorder A Questionnaire and Polysomnographic StudyDokument9 Seiten2007 Sleep in Children With Autistic Spectrum Disorder A Questionnaire and Polysomnographic StudypsicosmosNoch keine Bewertungen
- 2001 The First-Night Effect May Last More Than One NightDokument8 Seiten2001 The First-Night Effect May Last More Than One NightpsicosmosNoch keine Bewertungen
- Sleep and Wakefulness in Normal Preadolescent ChildrenDokument10 SeitenSleep and Wakefulness in Normal Preadolescent ChildrenpsicosmosNoch keine Bewertungen
- Vaccine Safety Study As An Interesting Case of "Over-Matching"Dokument14 SeitenVaccine Safety Study As An Interesting Case of "Over-Matching"psicosmosNoch keine Bewertungen
- Psychogenic Amnesia - A Malady of The Constricted Self SatniloiuDokument24 SeitenPsychogenic Amnesia - A Malady of The Constricted Self SatniloiupsicosmosNoch keine Bewertungen
- 1 s2.0 S1750946713000366 Main PDFDokument13 Seiten1 s2.0 S1750946713000366 Main PDFpsicosmosNoch keine Bewertungen
- Early Intervention of Autism: A Case For Floor Time ApproachDokument28 SeitenEarly Intervention of Autism: A Case For Floor Time ApproachpsicosmosNoch keine Bewertungen
- Clinical Research: Neuropsychological Assessment in Autism Spectrum Disorder and Related ConditionsDokument8 SeitenClinical Research: Neuropsychological Assessment in Autism Spectrum Disorder and Related ConditionspsicosmosNoch keine Bewertungen
- Research in Autism Spectrum Disorders: Li Yi, Yuebo Fan, Lisa Joseph, Dan Huang, Xueqin Wang, Jiao Li, Xiaobing ZouDokument7 SeitenResearch in Autism Spectrum Disorders: Li Yi, Yuebo Fan, Lisa Joseph, Dan Huang, Xueqin Wang, Jiao Li, Xiaobing ZoupsicosmosNoch keine Bewertungen
- Iran J Micro Bio 2014Dokument9 SeitenIran J Micro Bio 2014Eva Pa'e ONoch keine Bewertungen
- Genul StaphylococcusDokument28 SeitenGenul StaphylococcusAna Maria RusuNoch keine Bewertungen
- Sunway University College Monash University Foundation Year (MUFY) 2010Dokument24 SeitenSunway University College Monash University Foundation Year (MUFY) 2010Sunway UniversityNoch keine Bewertungen
- Daily Lesson Plan: 7e's ApproachDokument6 SeitenDaily Lesson Plan: 7e's ApproachSonny MagdadaroNoch keine Bewertungen
- Villacís2019 Article EfficacyOfPulsed-xenonUltravioDokument6 SeitenVillacís2019 Article EfficacyOfPulsed-xenonUltravioByron JimenezNoch keine Bewertungen
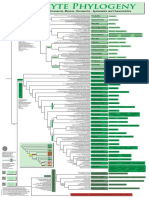
- Filogenia de Briofitos SensulatoDokument1 SeiteFilogenia de Briofitos SensulatoYuyitoS2714Noch keine Bewertungen
- Effects of Heat-Stress On Production in Dairy CattleDokument14 SeitenEffects of Heat-Stress On Production in Dairy CattleHussain Ahmad QadriNoch keine Bewertungen
- 8 13 PDFDokument7 Seiten8 13 PDFVillycia LNoch keine Bewertungen
- International Union For The Protection of New Varieties of PlantsDokument31 SeitenInternational Union For The Protection of New Varieties of PlantsBimo HariokusumoNoch keine Bewertungen
- Ethylhexylglycerin Safety ToxicologyDokument17 SeitenEthylhexylglycerin Safety Toxicologybfh83730Noch keine Bewertungen
- Wow Science Book-4Dokument52 SeitenWow Science Book-4Charu PawarNoch keine Bewertungen
- Post Harvest Tobacco Infestation ControlDokument160 SeitenPost Harvest Tobacco Infestation ControlKhosiKuneneNoch keine Bewertungen
- Biology The Essentials 2nd Edition by Marielle Hoefnagels ISBN Solution ManualDokument10 SeitenBiology The Essentials 2nd Edition by Marielle Hoefnagels ISBN Solution Manualmichael100% (25)
- CELL CYCLE PresentationDokument22 SeitenCELL CYCLE PresentationCalvin YusopNoch keine Bewertungen
- Biology 1990-2020 Al Ce Dse LQDokument238 SeitenBiology 1990-2020 Al Ce Dse LQ8kk-1Noch keine Bewertungen
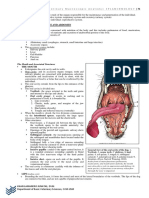
- Module 5 Anatomy I Splanchnology 1Dokument17 SeitenModule 5 Anatomy I Splanchnology 1neannaNoch keine Bewertungen
- Hellagrolip Nitrogenous Brochure enDokument2 SeitenHellagrolip Nitrogenous Brochure enBashir A. SialNoch keine Bewertungen
- 9780702072987-Book ChapterDokument2 Seiten9780702072987-Book ChaptervisiniNoch keine Bewertungen
- LipidDokument36 SeitenLipidSina AgataNoch keine Bewertungen
- Microbiomes Health and The Environment 1676562923Dokument733 SeitenMicrobiomes Health and The Environment 1676562923SagenxNoch keine Bewertungen
- 1,4 ButanediolDokument6 Seiten1,4 ButanediolVictor VikeneNoch keine Bewertungen
- NANDA Nursing Diagnosis List 2018-2020: Domain 1: Health PromotionDokument6 SeitenNANDA Nursing Diagnosis List 2018-2020: Domain 1: Health PromotionAngelaNoch keine Bewertungen
- Koi On Rearing Pond As ImmunostimulanDokument18 SeitenKoi On Rearing Pond As ImmunostimulanDodi SaputraNoch keine Bewertungen
- Mushrooms-Rich Preparations On Wound Healing: From Nutritional To Medicinal AttributesDokument9 SeitenMushrooms-Rich Preparations On Wound Healing: From Nutritional To Medicinal AttributesFilipe GonçalvesNoch keine Bewertungen
- Tutorial Nematoda 1 2019Dokument22 SeitenTutorial Nematoda 1 2019Andre HimawanNoch keine Bewertungen