Das könnte Ihnen auch gefallen
- Bowel Nosode MatrixDokument3 SeitenBowel Nosode MatrixAndrea Middleton92% (13)
- Spirometer Handbook NacaDokument24 SeitenSpirometer Handbook NacaebrycNoch keine Bewertungen
- Clockwork GenesDokument81 SeitenClockwork GenesTyler100% (2)
- The "Rail Sign": An Ultrasound Finding in Placenta Accreta Spectrum Indicating Deep Villous Invasion and Adverse OutcomesDokument17 SeitenThe "Rail Sign": An Ultrasound Finding in Placenta Accreta Spectrum Indicating Deep Villous Invasion and Adverse OutcomesJoseLuisNoch keine Bewertungen
- Pemberian Obat Pada AnakDokument19 SeitenPemberian Obat Pada AnakMuhamad RockystankiNoch keine Bewertungen
- Healing CodesDokument17 SeitenHealing CodesReiki Kiran95% (22)
- Cost - Effectiveness Analysis (CEA)Dokument9 SeitenCost - Effectiveness Analysis (CEA)Yeliana Hani RitongaNoch keine Bewertungen
- The Fundamental of Anti-Aging Medicine: Wimpie PangkahilaDokument44 SeitenThe Fundamental of Anti-Aging Medicine: Wimpie PangkahilajujuNoch keine Bewertungen
- The Analysis of Service Quality's Dimensional Based On Pasuraman Servqual Theory On JKN KIS Participant Satisfaction at Rolas Medika Clinic, JemberDokument9 SeitenThe Analysis of Service Quality's Dimensional Based On Pasuraman Servqual Theory On JKN KIS Participant Satisfaction at Rolas Medika Clinic, JemberInternational Journal of Innovative Science and Research TechnologyNoch keine Bewertungen
- PRP (Platelet Rich Plasma) Treatment in NepalDokument2 SeitenPRP (Platelet Rich Plasma) Treatment in NepalHealthy Choice ClinicNoch keine Bewertungen
- KP 1.1.3.4 Dan 1.1.3.6 Ebm PICODokument40 SeitenKP 1.1.3.4 Dan 1.1.3.6 Ebm PICOmuthia saniNoch keine Bewertungen
- Jadwal Operasi Senin, 5 Februari 2024Dokument1 SeiteJadwal Operasi Senin, 5 Februari 2024asandiNoch keine Bewertungen
- Vidas Brahms PCT Procalcitonin Brochure 1Dokument4 SeitenVidas Brahms PCT Procalcitonin Brochure 1swbartlinNoch keine Bewertungen
- Ascorbic Acid Injection, USPDokument4 SeitenAscorbic Acid Injection, USPYovie PrayektiNoch keine Bewertungen
- Male Pattern Baldness: A Survival GuideDokument11 SeitenMale Pattern Baldness: A Survival GuideMuhammad AliNoch keine Bewertungen
- Medication Safety RSDokument60 SeitenMedication Safety RSDeboraNainggolanNoch keine Bewertungen
- Data Laporan Pemakaian Obat Instalasi Farmasi Rumah Sakit QDokument4 SeitenData Laporan Pemakaian Obat Instalasi Farmasi Rumah Sakit QayuovianiNoch keine Bewertungen
- Baju RakorDokument32 SeitenBaju RakorDPK Rumkit Bhayangkara Tk INoch keine Bewertungen
- CDK Edisi CME-2 TH 2020 5 SKP PDFDokument24 SeitenCDK Edisi CME-2 TH 2020 5 SKP PDFnarinaaviNoch keine Bewertungen
- Kode TopografiDokument69 SeitenKode TopografiMUHAMMAD RASYID RNoch keine Bewertungen
- Test ID: PCT Procalcitonin, Serum: Useful ForDokument5 SeitenTest ID: PCT Procalcitonin, Serum: Useful ForI Nyoman Gede SemarajanaNoch keine Bewertungen
- Jurnal Polimer Fix PDFDokument5 SeitenJurnal Polimer Fix PDFSherly Marcia DevanaNoch keine Bewertungen
- Retinoids in TherapyDokument52 SeitenRetinoids in TherapyPanini PatankarNoch keine Bewertungen
- Far Mako Eko NomiDokument59 SeitenFar Mako Eko NomiLina LinuxNoch keine Bewertungen
- Penggunaan Obat Herbal Pada Pasien Kanker ServiksDokument9 SeitenPenggunaan Obat Herbal Pada Pasien Kanker ServiksislawatiNoch keine Bewertungen
- Final ANNOUNCEMENT 2019 Warna Orange EditDokument9 SeitenFinal ANNOUNCEMENT 2019 Warna Orange EditBimo Aryo TejoNoch keine Bewertungen
- Beban Kerja Perawat Terhadap Implementasi Patient Safety Di RuangDokument9 SeitenBeban Kerja Perawat Terhadap Implementasi Patient Safety Di RuangReno LogickNoch keine Bewertungen
- 12.naskah Publikasi PDFDokument25 Seiten12.naskah Publikasi PDFA'IkqNoch keine Bewertungen
- Vita Medika: Klinik Rawat InapDokument24 SeitenVita Medika: Klinik Rawat InapTri Anggun MayNoch keine Bewertungen
- Price List Alkes Pt. Carmella Gustavindo PERIODE JANUARY 2021 (Week 2)Dokument21 SeitenPrice List Alkes Pt. Carmella Gustavindo PERIODE JANUARY 2021 (Week 2)naelarizqiNoch keine Bewertungen
- Harga Ecatalog Reguler TERUMODokument4 SeitenHarga Ecatalog Reguler TERUMOrahayuNoch keine Bewertungen
- SINGKATANDokument26 SeitenSINGKATANOkto FanesaNoch keine Bewertungen
- ID Analisis Kebutuhan Tenaga Di Instalasi F PDFDokument9 SeitenID Analisis Kebutuhan Tenaga Di Instalasi F PDFYuanita PurnamiNoch keine Bewertungen
- Aceclofenac Efficacy and Safety Vs DiclofenacDokument6 SeitenAceclofenac Efficacy and Safety Vs DiclofenacpdelaxNoch keine Bewertungen
- Acne in The Adult: A. Ascenso and H. Cabral MarquesDokument10 SeitenAcne in The Adult: A. Ascenso and H. Cabral MarquesFrontiersNoch keine Bewertungen
- Guideline Kontrasepsi 1Dokument62 SeitenGuideline Kontrasepsi 1Laila Azzahrah SilitongaNoch keine Bewertungen
- Tabel ICD 10 English Indonesia LengkapDokument432 SeitenTabel ICD 10 English Indonesia LengkaptwindaNoch keine Bewertungen
- Drug Utilization Study (Dus)Dokument17 SeitenDrug Utilization Study (Dus)Arif Budiman100% (1)
- PPM Executive Development Syllabus 2021Dokument175 SeitenPPM Executive Development Syllabus 2021Miss ChessNoch keine Bewertungen
- Analisis Pelayanan Terapeutik Pada Pasien Tuberkulosis Resisten Obat Di Kota MedanDokument14 SeitenAnalisis Pelayanan Terapeutik Pada Pasien Tuberkulosis Resisten Obat Di Kota MedanwahyuNoch keine Bewertungen
- Hair Transplant Consent FormDokument7 SeitenHair Transplant Consent FormHossam Abd algawadNoch keine Bewertungen
- Arjati Daud PDFDokument9 SeitenArjati Daud PDFRizal AdiwangsaNoch keine Bewertungen
- Manajemen LabDokument42 SeitenManajemen LableniNoch keine Bewertungen
- Aspek Mikrobiologi Hepatitis Akut Berat Yang Belum DiketahuiDokument21 SeitenAspek Mikrobiologi Hepatitis Akut Berat Yang Belum DiketahuiSisiliana KristinNoch keine Bewertungen
- 2 Cost Utility AnalysisDokument29 Seiten2 Cost Utility Analysisapi-3723612Noch keine Bewertungen
- Intranatal Form FixDokument38 SeitenIntranatal Form FixChandra DewiNoch keine Bewertungen
- Implementasi Metode Kaizen Terhadap Kepuasan Pasien Di Instalasi Farmasi RuDokument6 SeitenImplementasi Metode Kaizen Terhadap Kepuasan Pasien Di Instalasi Farmasi RuEndai FajarNoch keine Bewertungen
- ManuskripDokument13 SeitenManuskripaamrii theartofracingNoch keine Bewertungen
- Genetic Disorders-1Dokument42 SeitenGenetic Disorders-1Bala UrmarNoch keine Bewertungen
- Eka Aryani 22010112110093 Lap KTI BAB 7Dokument36 SeitenEka Aryani 22010112110093 Lap KTI BAB 7Aldis Al FadjrinNoch keine Bewertungen
- NBSS AIA - Ivan Christian ChannelDokument128 SeitenNBSS AIA - Ivan Christian ChannelASTAGINA NAURAHNoch keine Bewertungen
- Tinpus FarmakoekonomiDokument6 SeitenTinpus FarmakoekonomiYesi Widya IswariNoch keine Bewertungen
- Materi RTDDokument16 SeitenMateri RTDJoenexusNoch keine Bewertungen
- Foundation Issues: Pharmacoeconomics: Principles, Methods, and ApplicationsDokument16 SeitenFoundation Issues: Pharmacoeconomics: Principles, Methods, and ApplicationsbencleeseNoch keine Bewertungen
- Dody Firmanda 2011 - Evidence-Based Medicine (EBM), Penilaian Teknologi Kesehatan (Health Technology Assessment/HTA) Dan Mutu Profesi MedisDokument18 SeitenDody Firmanda 2011 - Evidence-Based Medicine (EBM), Penilaian Teknologi Kesehatan (Health Technology Assessment/HTA) Dan Mutu Profesi MedisDody FirmandaNoch keine Bewertungen
- Pharmacogenomics: From Discovery to Clinical ImplementationVon EverandPharmacogenomics: From Discovery to Clinical ImplementationShowkat Ahmad GanieNoch keine Bewertungen
- Apoptosis: Apoptosis (From Ancient GreekDokument26 SeitenApoptosis: Apoptosis (From Ancient Greekuci kinaseNoch keine Bewertungen
- Extrinsic and Intrinsic Apoptosis Signal Pathway Review: Zhao HongmeiDokument20 SeitenExtrinsic and Intrinsic Apoptosis Signal Pathway Review: Zhao HongmeiAgustina PramudianingtiasNoch keine Bewertungen
- Apoptosis Programmed Cell Death and Its SignalsDokument11 SeitenApoptosis Programmed Cell Death and Its Signalsonur ateşNoch keine Bewertungen
- Apoptosis en Tejidos OralesDokument4 SeitenApoptosis en Tejidos Oralesangie catalina estupiñan mirandaNoch keine Bewertungen
- Apoptosis 5Dokument9 SeitenApoptosis 5Kamila Środa-PomianekNoch keine Bewertungen
- Necrotic Cell Death in Response To Oxidant Stress Involves The Activation of The Apoptogenic Caspase-8 Bid PathwayDokument8 SeitenNecrotic Cell Death in Response To Oxidant Stress Involves The Activation of The Apoptogenic Caspase-8 Bid PathwayalihamraghaniNoch keine Bewertungen
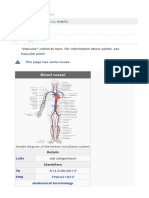
- Blood VesselDokument6 SeitenBlood VesselAnonymous VECoBcRSNoch keine Bewertungen
- Anne of ClevesDokument3 SeitenAnne of ClevesAnonymous VECoBcRSNoch keine Bewertungen
- BritishDokument3 SeitenBritishAnonymous VECoBcRSNoch keine Bewertungen
- Catherine ParrDokument3 SeitenCatherine ParrAnonymous VECoBcRSNoch keine Bewertungen
- Retinoblastoma Case Report Latest With ReferenceDokument29 SeitenRetinoblastoma Case Report Latest With ReferenceMasykur Ramadhan GaniNoch keine Bewertungen
- 1101 FullDokument8 Seiten1101 FullAprilia R. PermatasariNoch keine Bewertungen
- JanitaDokument2 SeitenJanitaapi-391376321Noch keine Bewertungen
- Nursing Care Plan For HypertensionDokument3 SeitenNursing Care Plan For HypertensionFranco Razon100% (2)
- Cardiovascular ExaminationDokument37 SeitenCardiovascular ExaminationmulatumeleseNoch keine Bewertungen
- Baby Friendly Hospital InitiativeDokument6 SeitenBaby Friendly Hospital InitiativeBinal JoshiNoch keine Bewertungen
- Case - IUFDDokument5 SeitenCase - IUFDMaris Sarline OpenianoNoch keine Bewertungen
- Cardiac Stress TestingDokument90 SeitenCardiac Stress TestingFrancesca Isabel BunyiNoch keine Bewertungen
- Building An NCD-ready Workforce: Technical Meeting ReportDokument48 SeitenBuilding An NCD-ready Workforce: Technical Meeting ReportnavenNoch keine Bewertungen
- UNEVERSITATEA DE STAT DE MEDICINA SI FARMACIE New PneumoDokument15 SeitenUNEVERSITATEA DE STAT DE MEDICINA SI FARMACIE New PneumoGalina LozovanuNoch keine Bewertungen
- MCQ Blok 11Dokument10 SeitenMCQ Blok 11Jefri SoniNoch keine Bewertungen
- Pitfalls PsmaDokument20 SeitenPitfalls Psmallana.mariabelenNoch keine Bewertungen
- Rectal Prolapse 5Dokument13 SeitenRectal Prolapse 5caliptra36Noch keine Bewertungen
- Resident's Page: Wood's Lamp Wood's Lamp Wood's Lamp Wood's Lamp Wood's LampDokument5 SeitenResident's Page: Wood's Lamp Wood's Lamp Wood's Lamp Wood's Lamp Wood's LampMovaliya GhanshyamNoch keine Bewertungen
- Radiation Oncology For Pediatric CNS Tumors - Mahajan & Paulino, 1E (2018)Dokument625 SeitenRadiation Oncology For Pediatric CNS Tumors - Mahajan & Paulino, 1E (2018)Bogdan Alexandru CristeiNoch keine Bewertungen
- Behcet DiseaseDokument6 SeitenBehcet DiseasejbahalkehNoch keine Bewertungen
- Intestinal TuberculosisDokument15 SeitenIntestinal Tuberculosisalfaz lakhaniNoch keine Bewertungen
- Managed Clinical Network For Stroke Stroke Unit Protocol/GuidelineDokument5 SeitenManaged Clinical Network For Stroke Stroke Unit Protocol/GuidelineRicky Cik KoharNoch keine Bewertungen
- Esophageal Disorder Part IIDokument27 SeitenEsophageal Disorder Part IIJuma AwarNoch keine Bewertungen
- Try Out NewDokument19 SeitenTry Out NewIke AdrianaNoch keine Bewertungen
- ICMM Health and Safety Performance Indicator DefinitionsDokument34 SeitenICMM Health and Safety Performance Indicator DefinitionsStakeholders360Noch keine Bewertungen
- LipomaDokument9 SeitenLipomashewasnot inrightmindNoch keine Bewertungen
- Calcium Metabolism: Preparation byDokument18 SeitenCalcium Metabolism: Preparation byAbhijeet KanjeNoch keine Bewertungen
- Abdominal ExamDokument7 SeitenAbdominal Examjoshy221196Noch keine Bewertungen
- Spinal Tumor StrongDokument45 SeitenSpinal Tumor StrongbettyNoch keine Bewertungen
- Ai Project ReportDokument12 SeitenAi Project ReportSaket SinghNoch keine Bewertungen
- Bentonite Material Safety Data Sheet PDFDokument6 SeitenBentonite Material Safety Data Sheet PDFWaquar AhmadNoch keine Bewertungen
- Dosages: Treatment of Influenza and Other DiseasesDokument6 SeitenDosages: Treatment of Influenza and Other Diseasesmartdot73490100% (8)