Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Cripple of Inishman - McDonaghDokument48 SeitenThe Cripple of Inishman - McDonaghelpeorerror100% (3)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- City Builder 03 - Entertainment PlacesDokument24 SeitenCity Builder 03 - Entertainment Placesskypalae100% (7)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Twilight TombDokument36 SeitenThe Twilight TombVictor Victorovich Khomyak100% (3)
- Science Quiz Bee EliminationDokument84 SeitenScience Quiz Bee EliminationDianne Diaz100% (7)
- Exploratory Latin AnimalsDokument15 SeitenExploratory Latin AnimalsKari Kristine Hoskins BarreraNoch keine Bewertungen
- Cleft Lip and Palate New ApproachDokument115 SeitenCleft Lip and Palate New ApproachsoorajNoch keine Bewertungen
- Fresh Fast Fabulous CookbookDokument140 SeitenFresh Fast Fabulous CookbookChase Ward100% (1)
- Violet Winspear The Awakening of AliceDokument197 SeitenViolet Winspear The Awakening of AliceSaumya Bhasin80% (5)
- Aries Kitten: Head & BodyDokument4 SeitenAries Kitten: Head & BodySelena KNoch keine Bewertungen
- Theories of Growth and DevelopmentDokument105 SeitenTheories of Growth and DevelopmentAjmal Najeeb50% (2)
- WBC BasicsDokument70 SeitenWBC BasicsZoe ZillaNoch keine Bewertungen
- Combining SentencesDokument21 SeitenCombining Sentencesfcfs1Noch keine Bewertungen
- Simplified New Energy WaysDokument36 SeitenSimplified New Energy WaysOBNoch keine Bewertungen
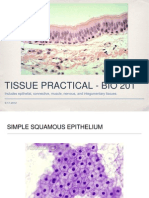
- Bio 201 - Tissue Practical (Powerpoint)Dokument53 SeitenBio 201 - Tissue Practical (Powerpoint)GretchenNoch keine Bewertungen
- Main IdeaDokument4 SeitenMain IdeaJohna Mae EtangNoch keine Bewertungen
- English 2 Periodical Test Quarter 4Dokument5 SeitenEnglish 2 Periodical Test Quarter 4April CelebradosNoch keine Bewertungen
- Skripsi Tanpa Bab PembahasanDokument79 SeitenSkripsi Tanpa Bab PembahasaniyelNoch keine Bewertungen
- Binding Capacity of Bull Spermatozoa ToDokument180 SeitenBinding Capacity of Bull Spermatozoa ToczumoffenNoch keine Bewertungen
- Visakha DairyDokument92 SeitenVisakha DairySrinivas Palukuri50% (2)
- Interesting Facts About English in No Particular Order...Dokument2 SeitenInteresting Facts About English in No Particular Order...Leticia PaniegoNoch keine Bewertungen
- Biological InteractionDokument17 SeitenBiological InteractionArchana SharmaNoch keine Bewertungen
- Care of Pt.Dokument6 SeitenCare of Pt.hobradorNoch keine Bewertungen
- Hematology Basics NewDokument32 SeitenHematology Basics Newkimbo23Noch keine Bewertungen
- Tiger and Tims Jungle AdventureDokument8 SeitenTiger and Tims Jungle AdventureTiger TimNoch keine Bewertungen
- The Cat of The Cane-BrakeDokument11 SeitenThe Cat of The Cane-Brakepaunero1222Noch keine Bewertungen
- AquariumDokument3 SeitenAquariumNafees AhmedNoch keine Bewertungen
- Hubungan Pemberian Imunisasi Dasar Dengan Tumbuh Kembang Pada Bayi (0 - 1 Tahun) Di Puskesmaskembes Kecamatan Tombulu Kabupaten MinahasaDokument10 SeitenHubungan Pemberian Imunisasi Dasar Dengan Tumbuh Kembang Pada Bayi (0 - 1 Tahun) Di Puskesmaskembes Kecamatan Tombulu Kabupaten MinahasatopanrkdNoch keine Bewertungen
- Abraham Blessing Cluster 988Dokument102 SeitenAbraham Blessing Cluster 988Becky ChiloveNoch keine Bewertungen
- Group I: DNA Viruses: PoxviridaeDokument3 SeitenGroup I: DNA Viruses: PoxviridaeharoononlineNoch keine Bewertungen
- PZOTEB0008E Dark TapestryDokument75 SeitenPZOTEB0008E Dark TapestryBraulioCharajaVargasNoch keine Bewertungen