Das könnte Ihnen auch gefallen
- Lopez Otin - 2016 Main PDFDokument20 SeitenLopez Otin - 2016 Main PDFDanilo RosaNoch keine Bewertungen
- Hormones and Reproduction of Vertebrates, Volume 4: BirdsVon EverandHormones and Reproduction of Vertebrates, Volume 4: BirdsNoch keine Bewertungen
- Chang2019 PDFDokument71 SeitenChang2019 PDFSolitario GeorgeNoch keine Bewertungen
- European Journal of PharmacologyDokument7 SeitenEuropean Journal of PharmacologyHartati HamziNoch keine Bewertungen
- Evolutionary Roots of Iodine and Thyroid HormonesDokument12 SeitenEvolutionary Roots of Iodine and Thyroid HormonesIsabella MejiaNoch keine Bewertungen
- Circadian Clock During Plant DevelopmentDokument9 SeitenCircadian Clock During Plant Development16. Erlangga Deanda Chandra SetyaNoch keine Bewertungen
- The Control of Body Size in Insects: H.F. NijhoutDokument9 SeitenThe Control of Body Size in Insects: H.F. NijhoutjocyeoNoch keine Bewertungen
- Microbiome Impact On Metabolism and Function of Sex, Thyroid, Growth and Parathyroid Hormones 2015Dokument13 SeitenMicrobiome Impact On Metabolism and Function of Sex, Thyroid, Growth and Parathyroid Hormones 2015Brenda FolkNoch keine Bewertungen
- Confounding Factors From Inducible Systems For Spatiotemporal Gene Expression RegulationDokument4 SeitenConfounding Factors From Inducible Systems For Spatiotemporal Gene Expression RegulationJessica Asitimbay ZuritaNoch keine Bewertungen
- A Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingDokument19 SeitenA Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingLautaro BelfioriNoch keine Bewertungen
- Dis. Model. Mech. 2010 Hortopan 144 8Dokument5 SeitenDis. Model. Mech. 2010 Hortopan 144 8Awanish MishraNoch keine Bewertungen
- Sun Et Al-2020-ElifeDokument76 SeitenSun Et Al-2020-ElifeScoots LimeNoch keine Bewertungen
- Review Environmental Epigenetics: A Role in Endocrine Disease?Dokument7 SeitenReview Environmental Epigenetics: A Role in Endocrine Disease?Najla MasturaNoch keine Bewertungen
- Specific Membrane Lipid Composition Is Important For Plasmodesmata Function in ArabidopsisDokument24 SeitenSpecific Membrane Lipid Composition Is Important For Plasmodesmata Function in ArabidopsisDary FarhanNoch keine Bewertungen
- Adverse Morphological Development in Embryonic Zebrafish Exposed To Environmental Concentrations of Contaminants Individually and in MixtureDokument13 SeitenAdverse Morphological Development in Embryonic Zebrafish Exposed To Environmental Concentrations of Contaminants Individually and in MixtureTiago TorresNoch keine Bewertungen
- Developmental Analysis of Lingo-1/Lern1 Protein Expression in The Mouse Brain: Interaction of Its Intracellular Domain With Myt1lDokument21 SeitenDevelopmental Analysis of Lingo-1/Lern1 Protein Expression in The Mouse Brain: Interaction of Its Intracellular Domain With Myt1lМария СавенкоNoch keine Bewertungen
- The Cell Biology of Aging: Race Diloreto and Coleen T. MurphyDokument8 SeitenThe Cell Biology of Aging: Race Diloreto and Coleen T. MurphyLeoberto Batista Pereira SobrinhoNoch keine Bewertungen
- Steroidogenesis and Its Regulation in Teleost-A ReviewDokument16 SeitenSteroidogenesis and Its Regulation in Teleost-A ReviewGeraldine Villamil JiménezNoch keine Bewertungen
- Bridget Project CompleteDokument70 SeitenBridget Project CompleteFavourNoch keine Bewertungen
- Journal of Pineal Research - 2017 - Mayo - Melatonin and Sirtuins A Not So Unexpected RelationshipDokument17 SeitenJournal of Pineal Research - 2017 - Mayo - Melatonin and Sirtuins A Not So Unexpected RelationshipBrenda AkemiNoch keine Bewertungen
- Mechanisms of Ageing and Development: Martin S. Denzel, Louis R. Lapierre, Hildegard I.D. MackDokument18 SeitenMechanisms of Ageing and Development: Martin S. Denzel, Louis R. Lapierre, Hildegard I.D. MackAle GuzmánNoch keine Bewertungen
- 1 s2.0 S0012160604004233 Main PDFDokument10 Seiten1 s2.0 S0012160604004233 Main PDFrocambolescas perthNoch keine Bewertungen
- Comparative Gene Expression Analysis of Genital TubercleDokument29 SeitenComparative Gene Expression Analysis of Genital TubercleAnonymous rprdjdFMNzNoch keine Bewertungen
- Insulin Analogs and Cancer: Laura Sciacca, Rosario Le Moli and Riccardo VigneriDokument9 SeitenInsulin Analogs and Cancer: Laura Sciacca, Rosario Le Moli and Riccardo VigneriSrinivas RajanalaNoch keine Bewertungen
- Dimorfismo SexualDokument21 SeitenDimorfismo SexualDiana CortesNoch keine Bewertungen
- Jara 2018Dokument54 SeitenJara 2018Torres Pineda OsvaldoNoch keine Bewertungen
- tmpB83A TMPDokument10 SeitentmpB83A TMPFrontiersNoch keine Bewertungen
- Mammalian Hibernation: Differential Gene Expression and Novel Application of Epigenetic ControlsDokument12 SeitenMammalian Hibernation: Differential Gene Expression and Novel Application of Epigenetic ControlsaufaaNoch keine Bewertungen
- Jacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentDokument11 SeitenJacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentCarly LeungNoch keine Bewertungen
- Autophagy-Independent FunctionsDokument18 SeitenAutophagy-Independent Functionselibb346Noch keine Bewertungen
- Molecular and Cellular Endocrinology: Endocrine Function in Naturally Long-Living Small MammalsDokument11 SeitenMolecular and Cellular Endocrinology: Endocrine Function in Naturally Long-Living Small MammalsJeremyPJmeNoch keine Bewertungen
- Impact of Endocrine Disruptor Chemicals in GynaecologyDokument14 SeitenImpact of Endocrine Disruptor Chemicals in GynaecologyVENTANABIOLOGICANoch keine Bewertungen
- Plcell v23 5 1985Dokument21 SeitenPlcell v23 5 1985Yulian NavaNoch keine Bewertungen
- Brain Aging and Regeneration After InjuriesDokument16 SeitenBrain Aging and Regeneration After InjuriesProsen21Noch keine Bewertungen
- Endocrine Disrupters - A Review of Some Sources, Effects, and Mechanisms of Actions On Behaviour and Neuroendocrine SystemsDokument28 SeitenEndocrine Disrupters - A Review of Some Sources, Effects, and Mechanisms of Actions On Behaviour and Neuroendocrine SystemsTiago TorresNoch keine Bewertungen
- Neuroglia in The Autistic Brain: Evidence From A Preclinical ModelDokument17 SeitenNeuroglia in The Autistic Brain: Evidence From A Preclinical ModelJose Alonso Aguilar ValeraNoch keine Bewertungen
- Chapters 1 5 References and AppendicesDokument88 SeitenChapters 1 5 References and AppendicesKai SanicoNoch keine Bewertungen
- Adiponectin Signaling and Function in Insulin Target TissuesDokument9 SeitenAdiponectin Signaling and Function in Insulin Target TissuesSanthiya KunasegaranNoch keine Bewertungen
- v1 457252 PDFDokument41 Seitenv1 457252 PDFAbhishek YadavNoch keine Bewertungen
- Different Effects of Anti-Sonic Hedgehog AntibodieDokument10 SeitenDifferent Effects of Anti-Sonic Hedgehog AntibodieVERONICA BLANCONoch keine Bewertungen
- 10 1016@j Tox 2012 11 001Dokument11 Seiten10 1016@j Tox 2012 11 001Muhammad FakhrizalNoch keine Bewertungen
- Thy-1 Modulates Neurological Cell-Cell and Cell-Matrix Interactions Through Multiple Molecular InteractionsDokument19 SeitenThy-1 Modulates Neurological Cell-Cell and Cell-Matrix Interactions Through Multiple Molecular InteractionsdupuytrenNoch keine Bewertungen
- The Brain DEDokument55 SeitenThe Brain DEpedroNoch keine Bewertungen
- British J Pharmacology - 2017 - Baronio - Embryonic Exposure To Valproic Acid Affects The Histaminergic System and TheDokument13 SeitenBritish J Pharmacology - 2017 - Baronio - Embryonic Exposure To Valproic Acid Affects The Histaminergic System and TheMadalina Andreea RobeaNoch keine Bewertungen
- Ery 153Dokument13 SeitenEry 153Miguel FriasNoch keine Bewertungen
- Uvod, Teme Seminara 2022Dokument22 SeitenUvod, Teme Seminara 2022tandifNoch keine Bewertungen
- 10.1007@s12031 015 0665 8Dokument11 Seiten10.1007@s12031 015 0665 8gina paola gina.paolaNoch keine Bewertungen
- Factors That Regulate Insulin Producing Cells and Their Output in DrosophilaDokument13 SeitenFactors That Regulate Insulin Producing Cells and Their Output in DrosophilaLautaro BelfioriNoch keine Bewertungen
- Ovarian ReserverDokument35 SeitenOvarian Reserverpramod singhNoch keine Bewertungen
- Differentiation: Jennifer E. Bestman, Krista D. Stackley, Jennifer J. Rahn, Tucker J. Williamson, Sherine S. L. ChanDokument19 SeitenDifferentiation: Jennifer E. Bestman, Krista D. Stackley, Jennifer J. Rahn, Tucker J. Williamson, Sherine S. L. ChanZeninJosepVásquezVillarNoch keine Bewertungen
- 1 s2.0 S0092867415001865 Main PDFDokument13 Seiten1 s2.0 S0092867415001865 Main PDFtlidiaNoch keine Bewertungen
- From Senescence-Promoting Genes To Anti-Aging PillDokument7 SeitenFrom Senescence-Promoting Genes To Anti-Aging PillKyler KingNoch keine Bewertungen
- Interplay Between Sugar and Hormone Signaling Pathways Modulate Floral Signal TransductionDokument12 SeitenInterplay Between Sugar and Hormone Signaling Pathways Modulate Floral Signal TransductionAfif BerlianNoch keine Bewertungen
- Glutamate Elicits Therapeutic Responses in Light-Induced Sleep-Deprived Zebrafish, Danio RerioDokument7 SeitenGlutamate Elicits Therapeutic Responses in Light-Induced Sleep-Deprived Zebrafish, Danio RerioDr Sharique AliNoch keine Bewertungen
- Seasonal Time Measurement During Reproduction: - ReviewDokument7 SeitenSeasonal Time Measurement During Reproduction: - ReviewRonie BrancoNoch keine Bewertungen
- Neuroinflamação e Doenças Mentais HHGHGDokument14 SeitenNeuroinflamação e Doenças Mentais HHGHGJuliana LimaNoch keine Bewertungen
- DRD4 and DAT1 in ADHD 04-12-09 No Reference SectionDokument51 SeitenDRD4 and DAT1 in ADHD 04-12-09 No Reference SectionJohanna Guzman0% (1)
- Role of Estrogen in Thyroid Function and Growth RegulationDokument8 SeitenRole of Estrogen in Thyroid Function and Growth RegulationLee Ming HinNoch keine Bewertungen
- Nilsson 2017Dokument18 SeitenNilsson 2017نذير الشرعبيNoch keine Bewertungen
- The Diagram Shows A Mother Fox and Her CubsDokument11 SeitenThe Diagram Shows A Mother Fox and Her CubsTansra KaurNoch keine Bewertungen
- Lect 2-Intro ContinuedDokument42 SeitenLect 2-Intro ContinuedSleeping BeautyNoch keine Bewertungen
- Zoology - Ijzr - Diversity of Spiders Around - Priya SharmaDokument8 SeitenZoology - Ijzr - Diversity of Spiders Around - Priya SharmaTJPRC PublicationsNoch keine Bewertungen
- Psarocolius - Decumanus - Crested Oropendola or CornbirdDokument6 SeitenPsarocolius - Decumanus - Crested Oropendola or CornbirdTheBoss 20Noch keine Bewertungen
- The Cerebellum-8 PDFDokument83 SeitenThe Cerebellum-8 PDFlovelots1234Noch keine Bewertungen
- Vocabulary: Animal QuizDokument1 SeiteVocabulary: Animal QuizMariciell Larissa Gonzales TiconaNoch keine Bewertungen
- Student's Book: Kim MilneDokument17 SeitenStudent's Book: Kim MilneЕлизавета БогоявленскаяNoch keine Bewertungen
- ALPHABET-activity Worksheets For KinderDokument84 SeitenALPHABET-activity Worksheets For KinderLeah Paulyn Bascos Bergonio-LozadaNoch keine Bewertungen
- Captive Care of North American Water TurtlesDokument160 SeitenCaptive Care of North American Water TurtlesAnthony Lopez SahagunNoch keine Bewertungen
- Handball Performance Physiological Considerations & Practical Approach For Training MetabolicDokument13 SeitenHandball Performance Physiological Considerations & Practical Approach For Training Metabolicsanta3Noch keine Bewertungen
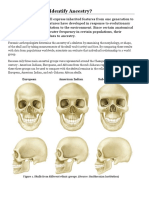
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanDokument5 SeitenActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyNoch keine Bewertungen
- Unit 4 Cell CommunicationDokument74 SeitenUnit 4 Cell CommunicationBerryNoch keine Bewertungen
- DAT QuizletDokument78 SeitenDAT QuizletJihee YoonNoch keine Bewertungen
- 14 - Scalp (FF)Dokument27 Seiten14 - Scalp (FF)checkmateNoch keine Bewertungen
- Biogeographical Regions PDFDokument7 SeitenBiogeographical Regions PDFmanoj_rkl_07100% (1)
- GI Tract Histology SummaryDokument5 SeitenGI Tract Histology SummaryFluffy_iceNoch keine Bewertungen
- Three Essays On The Dinosaurs - Immanuel VelikovskyDokument6 SeitenThree Essays On The Dinosaurs - Immanuel VelikovskyBright StarrNoch keine Bewertungen
- Land AnimalsDokument52 SeitenLand AnimalsVinuta AilNoch keine Bewertungen
- NURSAKINAH NAJWAH BT SUHAIMI (Dissection and Observation of Plant Reproductive Organs)Dokument9 SeitenNURSAKINAH NAJWAH BT SUHAIMI (Dissection and Observation of Plant Reproductive Organs)Nursakinah NajwahNoch keine Bewertungen
- đề thi preDokument14 Seitenđề thi prePham congNoch keine Bewertungen
- English 3RD TestDokument3 SeitenEnglish 3RD TestMichael CalesajrNoch keine Bewertungen
- Raz lr28 Birdbeaks CLRDokument8 SeitenRaz lr28 Birdbeaks CLRapi-290807931Noch keine Bewertungen
- Distinguishing Features of Anthropology - Characteristics 1-5 6.78 What Are The3 Unique Approaches of AnthropologyDokument2 SeitenDistinguishing Features of Anthropology - Characteristics 1-5 6.78 What Are The3 Unique Approaches of AnthropologymatrixtrinityNoch keine Bewertungen
- Eukaryotes and ProkaryotesDokument31 SeitenEukaryotes and ProkaryotesAysha MohideenNoch keine Bewertungen
- English Questionnaire For Final Exam. January 20th 2022Dokument4 SeitenEnglish Questionnaire For Final Exam. January 20th 2022Daniel CabreraNoch keine Bewertungen
- 2008 Villanueva & Norman 2008 With Colour PlatesDokument105 Seiten2008 Villanueva & Norman 2008 With Colour PlatesEstefany Lopez RipollNoch keine Bewertungen
- Structure of Woody Plants: Notes Made From Mauseth and RavenDokument13 SeitenStructure of Woody Plants: Notes Made From Mauseth and RavenhomamunfatNoch keine Bewertungen
- Listening Test #9Dokument3 SeitenListening Test #9personalmail_20011078Noch keine Bewertungen
- Lecture 2 - HistologyDokument7 SeitenLecture 2 - HistologyMaddie HallNoch keine Bewertungen
- Exploring Animal Life Cycles: Ms. Mckeel & Ms. Messina 2nd GradeDokument15 SeitenExploring Animal Life Cycles: Ms. Mckeel & Ms. Messina 2nd Gradeapi-302056293Noch keine Bewertungen
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseVon EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseBewertung: 4.5 von 5 Sternen4.5/5 (52)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldVon EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldBewertung: 4 von 5 Sternen4/5 (597)
- Why We Die: The New Science of Aging and the Quest for ImmortalityVon EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityBewertung: 4 von 5 Sternen4/5 (5)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsVon EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsBewertung: 4.5 von 5 Sternen4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisVon EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisBewertung: 3.5 von 5 Sternen3.5/5 (2)
- Gut: the new and revised Sunday Times bestsellerVon EverandGut: the new and revised Sunday Times bestsellerBewertung: 4 von 5 Sternen4/5 (393)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomVon EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomBewertung: 4 von 5 Sternen4/5 (216)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessVon Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessBewertung: 4 von 5 Sternen4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceVon EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceBewertung: 4.5 von 5 Sternen4.5/5 (517)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeVon EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNoch keine Bewertungen
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Von EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Bewertung: 4 von 5 Sternen4/5 (411)
- Tales from Both Sides of the Brain: A Life in NeuroscienceVon EverandTales from Both Sides of the Brain: A Life in NeuroscienceBewertung: 3 von 5 Sternen3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesVon EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesBewertung: 4.5 von 5 Sternen4.5/5 (397)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedVon EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedBewertung: 4 von 5 Sternen4/5 (11)
- Seven and a Half Lessons About the BrainVon EverandSeven and a Half Lessons About the BrainBewertung: 4 von 5 Sternen4/5 (110)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionVon EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionBewertung: 4 von 5 Sternen4/5 (812)
- Who's in Charge?: Free Will and the Science of the BrainVon EverandWho's in Charge?: Free Will and the Science of the BrainBewertung: 4 von 5 Sternen4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorVon EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNoch keine Bewertungen
- Lymph & Longevity: The Untapped Secret to HealthVon EverandLymph & Longevity: The Untapped Secret to HealthBewertung: 4.5 von 5 Sternen4.5/5 (13)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildVon EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildBewertung: 4.5 von 5 Sternen4.5/5 (44)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindVon EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindBewertung: 4.5 von 5 Sternen4.5/5 (93)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemVon EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemBewertung: 4.5 von 5 Sternen4.5/5 (115)
- Why We Sleep: Unlocking the Power of Sleep and DreamsVon EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsBewertung: 4.5 von 5 Sternen4.5/5 (2083)
- Good Without God: What a Billion Nonreligious People Do BelieveVon EverandGood Without God: What a Billion Nonreligious People Do BelieveBewertung: 4 von 5 Sternen4/5 (66)