Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1091)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Diabetes Mellitus Drug ChartDokument3 SeitenDiabetes Mellitus Drug Chartlui.stephanie1751100% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- SemaglutideDokument95 SeitenSemaglutideShigma Putra Mahaley100% (1)
- Chapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasDokument17 SeitenChapter 60 - Insulin, Oral Hypoglycaemic Agents, and The Pharmacology of The Endocrine PancreasGiliana P. García AcevedoNoch keine Bewertungen
- Juan Carlos Vsim Prep 3Dokument5 SeitenJuan Carlos Vsim Prep 3Michelle Pinkhasova100% (3)
- BMJ 2022 074068.fullDokument13 SeitenBMJ 2022 074068.fullANoch keine Bewertungen
- CardioVascular and Endocrin ExercisesDokument6 SeitenCardioVascular and Endocrin ExercisesBogartNoch keine Bewertungen
- Diabetes Mellitus Type 1 and 2Dokument11 SeitenDiabetes Mellitus Type 1 and 2Ibrar AliNoch keine Bewertungen
- Case Report - Diabetes MellitusDokument19 SeitenCase Report - Diabetes MellitusAnggitafhNoch keine Bewertungen
- Extraction, Isolation and Structural Elucidation of Flavonoid FromDokument10 SeitenExtraction, Isolation and Structural Elucidation of Flavonoid FromYuliet SusantoNoch keine Bewertungen
- Analytical Methods: Accepted ManuscriptDokument39 SeitenAnalytical Methods: Accepted ManuscriptYuliet SusantoNoch keine Bewertungen
- 225 2017-CJFSDokument8 Seiten225 2017-CJFSYuliet SusantoNoch keine Bewertungen
- Evaluation of Hypoglycemic and Hypolipidemic Activity of Methanol Extract of Brassica OleraceaDokument6 SeitenEvaluation of Hypoglycemic and Hypolipidemic Activity of Methanol Extract of Brassica OleraceaYuliet SusantoNoch keine Bewertungen
- Antioxidant and Hypoglycemic Activities of Extract and Fractions of Rambutan Seeds (Nephelium Lappaceum L.)Dokument6 SeitenAntioxidant and Hypoglycemic Activities of Extract and Fractions of Rambutan Seeds (Nephelium Lappaceum L.)Yuliet SusantoNoch keine Bewertungen
- PharmacognJ 10 71 - 2 PDFDokument6 SeitenPharmacognJ 10 71 - 2 PDFYuliet SusantoNoch keine Bewertungen
- Study of The Antidiabetic Activity of Punica Granatum L. Fruits Aqueous Extract On The Alloxan-Diabetic Wistar RatsDokument11 SeitenStudy of The Antidiabetic Activity of Punica Granatum L. Fruits Aqueous Extract On The Alloxan-Diabetic Wistar RatsYuliet SusantoNoch keine Bewertungen
- Sciencedirect: N.K. Achi, O.C. Ohaeri, I.I. Ijeh, C. EleazuDokument8 SeitenSciencedirect: N.K. Achi, O.C. Ohaeri, I.I. Ijeh, C. EleazuYuliet SusantoNoch keine Bewertungen
- Accepted ManuscriptDokument22 SeitenAccepted ManuscriptYuliet SusantoNoch keine Bewertungen
- 1 s2.0 S2225411016302140 MainDokument10 Seiten1 s2.0 S2225411016302140 MainYuliet SusantoNoch keine Bewertungen
- The Hospital Management of Hypoglycaemia in Adults With Diabetes Mellitus 4th EditionDokument40 SeitenThe Hospital Management of Hypoglycaemia in Adults With Diabetes Mellitus 4th EditionAmalia SholihahNoch keine Bewertungen
- En Diamicron MR CiDokument4 SeitenEn Diamicron MR CiDewie DeAlove AnggarRiniNoch keine Bewertungen
- Sulfonylurea Pharmacology Updates 2020Dokument3 SeitenSulfonylurea Pharmacology Updates 2020AHMED TANJIMUL ISLAMNoch keine Bewertungen
- Oral Pharmacologic Treatment of Type 2 Diabetes Mellitus: A Clinical Practice Guideline Update From The American College of PhysiciansDokument20 SeitenOral Pharmacologic Treatment of Type 2 Diabetes Mellitus: A Clinical Practice Guideline Update From The American College of PhysiciansRoberto López MataNoch keine Bewertungen
- Diabetes - Oral KnowledgeDokument19 SeitenDiabetes - Oral KnowledgeKSeegurNoch keine Bewertungen
- Synthesis and Pharmacological Evaluation of Some Novel Potent Type Ii Antidiabetic AgentsDokument10 SeitenSynthesis and Pharmacological Evaluation of Some Novel Potent Type Ii Antidiabetic AgentsmetalnonmetalNoch keine Bewertungen
- Mounjaro UspiDokument19 SeitenMounjaro UspiayaznptiNoch keine Bewertungen
- Efficacy and Safety of Biosimilar Glargine-IMGDokument8 SeitenEfficacy and Safety of Biosimilar Glargine-IMGSiddiq MohammedNoch keine Bewertungen
- Repaglinide LDokument19 SeitenRepaglinide LbigbullsrjNoch keine Bewertungen
- GlipizideDokument3 SeitenGlipizideapi-3797941100% (1)
- Evaluating The Effects of Insulin, Metformin and Glibenclamide On The Pups' Prefrontal Cortex and Oxidative Stress Markers of Streptozotocin-Induced Diabetic Pregnant RatsDokument10 SeitenEvaluating The Effects of Insulin, Metformin and Glibenclamide On The Pups' Prefrontal Cortex and Oxidative Stress Markers of Streptozotocin-Induced Diabetic Pregnant RatsMediterr J Pharm Pharm SciNoch keine Bewertungen
- Diabetes TreatmentDokument14 SeitenDiabetes TreatmentmusimsemiNoch keine Bewertungen
- Diabetes Mellitus in ElderlyDokument11 SeitenDiabetes Mellitus in ElderlyTheodora IonescuNoch keine Bewertungen
- Diabetes PpsDokument60 SeitenDiabetes PpsAzhi Ima AwufiNoch keine Bewertungen
- Obat-Obat Baru 2010Dokument262 SeitenObat-Obat Baru 2010UdtjeVanDerJeykNoch keine Bewertungen
- DM and GlucoseDokument1 SeiteDM and GlucoseibunaraNoch keine Bewertungen
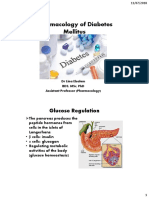
- Pharmacology of Diabetes Mellitus: Glucose RegulationDokument33 SeitenPharmacology of Diabetes Mellitus: Glucose RegulationAbdullah RawashdehNoch keine Bewertungen
- IT 10 - Farmakologi Obat - Obat Di Bidang Endokrin - TEODokument34 SeitenIT 10 - Farmakologi Obat - Obat Di Bidang Endokrin - TEOtattosssNoch keine Bewertungen
- Meglitinides ProjectDokument10 SeitenMeglitinides Projectarmaan kaltaNoch keine Bewertungen
- Pioglitazone A Review of Its Use in Type 2.51Dokument26 SeitenPioglitazone A Review of Its Use in Type 2.51Nurhadi EffendyNoch keine Bewertungen
- Anti Diabetik Oral Dan InsulinDokument97 SeitenAnti Diabetik Oral Dan InsulinunknownNoch keine Bewertungen
- JalraDokument3 SeitenJalraPrachi Gupta100% (1)