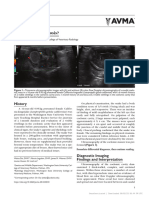
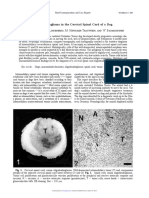
Das könnte Ihnen auch gefallen
- Barber 2004Dokument18 SeitenBarber 2004camilaNoch keine Bewertungen
- Amphibian OncologyDokument23 SeitenAmphibian OncologyAli JaraNoch keine Bewertungen
- Comparative Clinical PathologyDokument5 SeitenComparative Clinical PathologyErin RiosNoch keine Bewertungen
- Organos LinfoidesDokument13 SeitenOrganos LinfoidesLina JohananaNoch keine Bewertungen
- On June 30, 2019 Downloaded From Published Online: 1 March, 1912 - Supp InfoDokument20 SeitenOn June 30, 2019 Downloaded From Published Online: 1 March, 1912 - Supp InfoAgungBudiPamungkasNoch keine Bewertungen
- The Lymphoid System: A Review of Species DifferencesDokument13 SeitenThe Lymphoid System: A Review of Species DifferencesValeria Garces FlorezNoch keine Bewertungen
- Regulation of Parasite PopulationsVon EverandRegulation of Parasite PopulationsGerald EschNoch keine Bewertungen
- Craniopharyngiomas: Niki Karavitaki, Simon Cudlip, Christopher B. T. Adams, and John A. H. WassDokument27 SeitenCraniopharyngiomas: Niki Karavitaki, Simon Cudlip, Christopher B. T. Adams, and John A. H. Wassخالد الشيبريNoch keine Bewertungen
- JZWM Jan 2010Dokument7 SeitenJZWM Jan 2010llcoffeeNoch keine Bewertungen
- A Compilation of Cellular and Tissue Xenotransplantation Studies (1838-2022): A Promising Approach for the Treatment of DiseasesVon EverandA Compilation of Cellular and Tissue Xenotransplantation Studies (1838-2022): A Promising Approach for the Treatment of DiseasesNoch keine Bewertungen
- Vet Radiology Ultrasound - 2023 - MacLeod - Ultrasonographic Appearance of Gallbladder Neoplasia in 14 Dogs and 1 CatDokument9 SeitenVet Radiology Ultrasound - 2023 - MacLeod - Ultrasonographic Appearance of Gallbladder Neoplasia in 14 Dogs and 1 CatnerakyokNoch keine Bewertungen
- 2022 - Left Shift and Toxic Change in Heterophils and Neutrophils of Non Mammalian-Stacy Et Al.Dokument27 Seiten2022 - Left Shift and Toxic Change in Heterophils and Neutrophils of Non Mammalian-Stacy Et Al.Rachel PulcherioNoch keine Bewertungen
- Left Shift and Toxic Change in Heterophils and Neutrophils Ofnon Mammalian Vertebrates: A Comparative Review, Imageatlas, and Practical ConsiderationsDokument27 SeitenLeft Shift and Toxic Change in Heterophils and Neutrophils Ofnon Mammalian Vertebrates: A Comparative Review, Imageatlas, and Practical ConsiderationsAkın SeverNoch keine Bewertungen
- Jaaha 1998 34 2Dokument78 SeitenJaaha 1998 34 2templarioNoch keine Bewertungen
- 1 - 2009 - Tulio Et Al. (2009) Paraparesia Espástica e Hiperreflexia em Um Bovino Associada A Carcinoma de Células Escamosas, Relato de CasoDokument7 Seiten1 - 2009 - Tulio Et Al. (2009) Paraparesia Espástica e Hiperreflexia em Um Bovino Associada A Carcinoma de Células Escamosas, Relato de CasoRoney ZimpelNoch keine Bewertungen
- J Toxicol Pathol 2017Dokument18 SeitenJ Toxicol Pathol 2017Lina JohananaNoch keine Bewertungen
- Em Pathology em 50 Years On - 2017 - PatholDokument2 SeitenEm Pathology em 50 Years On - 2017 - PatholHamed AhmedNoch keine Bewertungen
- Modpathol 201783Dokument14 SeitenModpathol 201783Tian Nopita SariNoch keine Bewertungen
- Invasive Epithelial Mesothelioma in A Dog: F. R, B. B, K. R, E. S, D. BDokument5 SeitenInvasive Epithelial Mesothelioma in A Dog: F. R, B. B, K. R, E. S, D. BjamesyuNoch keine Bewertungen
- Initial Impact of The Sequencing of The Human GenomeDokument11 SeitenInitial Impact of The Sequencing of The Human GenomeMariano PerezNoch keine Bewertungen
- Feline Gastrointestinal Eosinofílica Fibroplasia EsclerosanteDokument8 SeitenFeline Gastrointestinal Eosinofílica Fibroplasia EsclerosanteCarlos Alberto Chaves VelasquezNoch keine Bewertungen
- Javma-Javma 20 04 0230Dokument3 SeitenJavma-Javma 20 04 0230Fiorella YavarNoch keine Bewertungen
- Tumours: Xxi. of Bones and JointsDokument18 SeitenTumours: Xxi. of Bones and JointsNeha PalsikarNoch keine Bewertungen
- Near-Infrared Raman Spectroscopy For TheDokument12 SeitenNear-Infrared Raman Spectroscopy For TheAlberto Carlos Espinosa GaravitoNoch keine Bewertungen
- RPB14150079015Dokument5 SeitenRPB14150079015Ijupbs IjupbsNoch keine Bewertungen
- 2022 Linf (AticoDokument12 Seiten2022 Linf (AticoJessica LaraNoch keine Bewertungen
- Guimier Et Al. (2015) PMID 26437028 - 0Dokument14 SeitenGuimier Et Al. (2015) PMID 26437028 - 0Filip MilošićNoch keine Bewertungen
- Oligodendroglioma in The Cervical Spinal Cord of A Dog: T. M, A. M - L, M. H - T, W. BDokument3 SeitenOligodendroglioma in The Cervical Spinal Cord of A Dog: T. M, A. M - L, M. H - T, W. BSamir BazanNoch keine Bewertungen
- Cytogenetics of Cells in CultureVon EverandCytogenetics of Cells in CultureR. J. C. HarrisNoch keine Bewertungen
- Cancer An Old Disease A New Disease or Something in BetweenDokument6 SeitenCancer An Old Disease A New Disease or Something in BetweenPaula VeigaNoch keine Bewertungen
- Insertional Mutagenesis Identifies Multiple Networks of Cooperating Genes Driving Intestinal TumorigenesisDokument10 SeitenInsertional Mutagenesis Identifies Multiple Networks of Cooperating Genes Driving Intestinal Tumorigenesistalha saleemNoch keine Bewertungen
- Seminoma Metastático: Descripción Histomorfológica: Metastatic Seminoma: Histomorphologic DescriptionDokument5 SeitenSeminoma Metastático: Descripción Histomorfológica: Metastatic Seminoma: Histomorphologic DescriptionEisson PitreNoch keine Bewertungen
- Haematology - An Illustrated Colour Text, 4th Edition - 2Dokument132 SeitenHaematology - An Illustrated Colour Text, 4th Edition - 2蔡季霖100% (1)
- Module 1 - CytogeneticsDokument5 SeitenModule 1 - CytogeneticsNursida LadjalawieNoch keine Bewertungen
- Canine Perineal TumoursDokument11 SeitenCanine Perineal TumoursSUSANA SAM RODRIGUEZNoch keine Bewertungen
- Thesis Hematopoietic Stem CellsDokument5 SeitenThesis Hematopoietic Stem Cellstunwpmzcf100% (1)
- Patogenesis Pleomorphic AdenomaDokument6 SeitenPatogenesis Pleomorphic AdenomaYeni PuspitasariNoch keine Bewertungen
- Aneuploidy-Induced Delaminating Cells Drive Tumorigenesis in Drosophila EpitheliaDokument6 SeitenAneuploidy-Induced Delaminating Cells Drive Tumorigenesis in Drosophila EpitheliaPierre RICHARD-PARPAILLONNoch keine Bewertungen
- Metastatic Extramedullary Plasmacytoma in A Canine 75186inglesDokument6 SeitenMetastatic Extramedullary Plasmacytoma in A Canine 75186inglesEzequiel Davi Dos SantosNoch keine Bewertungen
- Cell Movement and Neoplasia: Proceedings of the Annual Meeting of the Cell Tissue and Organ Culture Study Group, Held at the Janssen Research Foundation, Beerse, Belgium, May 1979Von EverandCell Movement and Neoplasia: Proceedings of the Annual Meeting of the Cell Tissue and Organ Culture Study Group, Held at the Janssen Research Foundation, Beerse, Belgium, May 1979M. De BrabanderBewertung: 5 von 5 Sternen5/5 (1)
- Chondro-Osseous Respiratory Epithelial Adenomatoid Hamartomas in 3 DogsDokument4 SeitenChondro-Osseous Respiratory Epithelial Adenomatoid Hamartomas in 3 DogsfisheirNoch keine Bewertungen
- Fidler 2001Dokument14 SeitenFidler 2001Claudia BrînzaNoch keine Bewertungen
- EScholarship UC Item 7s43p5x8Dokument4 SeitenEScholarship UC Item 7s43p5x8Hafich ErnandaNoch keine Bewertungen
- Third Eyelid Gland Neoplasms of Dogs and CatsDokument6 SeitenThird Eyelid Gland Neoplasms of Dogs and CatsMicheleBerselliNoch keine Bewertungen
- Thesis On Embryonic Stem CellsDokument10 SeitenThesis On Embryonic Stem CellsGina Rizzo100% (2)
- CADAE1405CDokument12 SeitenCADAE1405CMuhammad RiduanNoch keine Bewertungen
- Hormones and Reproduction of Vertebrates, Volume 3: ReptilesVon EverandHormones and Reproduction of Vertebrates, Volume 3: ReptilesNoch keine Bewertungen
- Solving the riddle of cancer: new genetic approaches to treatmentVon EverandSolving the riddle of cancer: new genetic approaches to treatmentNoch keine Bewertungen
- Necropsy - Killer Whale TakuDokument5 SeitenNecropsy - Killer Whale TakuThe Orca Project Corp100% (3)
- Histogenesis of Salivary Gland NeoplasmsDokument18 SeitenHistogenesis of Salivary Gland Neoplasmsporkodi sudhaNoch keine Bewertungen
- Congenital Cystic Disease of The Liver, Pancreas, and Kidney in A Nubian GoatDokument3 SeitenCongenital Cystic Disease of The Liver, Pancreas, and Kidney in A Nubian GoatEma SomantriNoch keine Bewertungen
- Establishment and CharacterizationDokument8 SeitenEstablishment and CharacterizationJosué Zuriel OrtizNoch keine Bewertungen
- Human EvolutionDokument374 SeitenHuman EvolutionMirabella Astra100% (10)
- tmp53 TMPDokument3 Seitentmp53 TMPFrontiersNoch keine Bewertungen
- Warthin Tumor of The Oropharyngeal Minor Salivary GlandDokument5 SeitenWarthin Tumor of The Oropharyngeal Minor Salivary GlandChairul LatiefNoch keine Bewertungen
- 6703 23736 1 PBDokument3 Seiten6703 23736 1 PBReski Harlianty HarliNoch keine Bewertungen
- Cancer Biology, a Study of Cancer Pathogenesis: How to Prevent Cancer and DiseasesVon EverandCancer Biology, a Study of Cancer Pathogenesis: How to Prevent Cancer and DiseasesNoch keine Bewertungen
- Robin's HistopathDokument6 SeitenRobin's HistopathKennedy DelrosarioNoch keine Bewertungen
- Intramedullary Spinal Cord TumorsDokument7 SeitenIntramedullary Spinal Cord TumorsmutalimNoch keine Bewertungen
- A 3 Table of Surgical Procedures TOSP With Minor Surgical Procedures MSPs 1Dokument94 SeitenA 3 Table of Surgical Procedures TOSP With Minor Surgical Procedures MSPs 1Chee Yung NgNoch keine Bewertungen
- Skin Tumor AgusDokument54 SeitenSkin Tumor AgusHafida AuliaristaNoch keine Bewertungen
- Endocrine ClarkDokument834 SeitenEndocrine ClarkKhaled AbdelgalelNoch keine Bewertungen
- Cancerul Si NeoplaziaDokument28 SeitenCancerul Si NeoplaziaBogdan SoltuzuNoch keine Bewertungen
- Brain Tumors - Classifications, Symptoms, Diagnosis and TreatmentsDokument10 SeitenBrain Tumors - Classifications, Symptoms, Diagnosis and TreatmentsshamimNoch keine Bewertungen
- Neoplasia HW (ANDRADE, Jhayber Jet L.) PDFDokument12 SeitenNeoplasia HW (ANDRADE, Jhayber Jet L.) PDFJhayber AndradeNoch keine Bewertungen
- All Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-MullaDokument45 SeitenAll Information Is On Websit:: Soft Tissue Tumours by Dr. Fahd Al-Mullariskhapangestika100% (1)
- Logistic Regression: Gunjan Bharadwaj Assistant Professor Dept of CEADokument42 SeitenLogistic Regression: Gunjan Bharadwaj Assistant Professor Dept of CEARahul Singh100% (1)
- Neoplasia NotesDokument10 SeitenNeoplasia NotesCharlene Fernández100% (4)
- Tumors: Benign and Malignant: Cancer Therapy & OncologyDokument3 SeitenTumors: Benign and Malignant: Cancer Therapy & OncologyMaria ArifahNoch keine Bewertungen
- Seminar 1Dokument41 SeitenSeminar 1Aprillia Dian PertiwiNoch keine Bewertungen
- IOTA - Simple - Rules - Susanne - Johnson 2 PDFDokument76 SeitenIOTA - Simple - Rules - Susanne - Johnson 2 PDFBenyamin Rakhmatsyah TitaleyNoch keine Bewertungen
- Adenoma Carcinoma SequenceDokument27 SeitenAdenoma Carcinoma SequenceKhurshedul AlamNoch keine Bewertungen
- Cellular AberrationsDokument94 SeitenCellular AberrationsKatherineCentenoIlaganRNNoch keine Bewertungen
- Brain Tumor Detection Using CNNDokument10 SeitenBrain Tumor Detection Using CNNIJRASETPublicationsNoch keine Bewertungen
- Pathoma Study OutlineDokument19 SeitenPathoma Study OutlineJustine Val RevalidaNoch keine Bewertungen
- NeoplasiaDokument78 SeitenNeoplasiageetikaNoch keine Bewertungen
- Oncologic NursingDokument132 SeitenOncologic Nursingɹǝʍdןnos100% (10)
- Diagnostic Imaging - Abdomen PDFDokument1.017 SeitenDiagnostic Imaging - Abdomen PDFLucian Vintila67% (3)
- Hemangioma - Google SearchDokument1 SeiteHemangioma - Google SearchhapidarrenNoch keine Bewertungen
- Cysts of Pancreas RADIOLOGYDokument20 SeitenCysts of Pancreas RADIOLOGYmhany12345Noch keine Bewertungen
- NCM 41118L Final Quiz 3Dokument13 SeitenNCM 41118L Final Quiz 3Matelyn OargaNoch keine Bewertungen
- Derm PathologyDokument21 SeitenDerm PathologyMaria GeorgeNoch keine Bewertungen
- Jabot 2009Dokument5 SeitenJabot 2009eric suNoch keine Bewertungen
- Appendigeal TumoursDokument3 SeitenAppendigeal TumourspritanuNoch keine Bewertungen
- Pathology Of: NeoplasiaDokument23 SeitenPathology Of: NeoplasiaBryan de GuzmanNoch keine Bewertungen
- Chapter 6 NeoplasiaDokument24 SeitenChapter 6 Neoplasiahenna patelNoch keine Bewertungen
- Case Report: A Case Report of Aggressive Chronic Rhinosinusitis With Nasal Polyps Mimicking Sinonasal MalignancyDokument6 SeitenCase Report: A Case Report of Aggressive Chronic Rhinosinusitis With Nasal Polyps Mimicking Sinonasal MalignancyRegina Dwindarti DarostyNoch keine Bewertungen