Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- William H. Soine - Clandestine Drug SynthesisDokument34 SeitenWilliam H. Soine - Clandestine Drug SynthesisAnonymous Gb9EyW83% (6)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Recreational Drug Discovery: Natural Products As Lead Structures For The Synthesis of Smart DrugsDokument26 SeitenRecreational Drug Discovery: Natural Products As Lead Structures For The Synthesis of Smart DrugsStiven DestNoch keine Bewertungen
- Drugs For Parkinson's DiseaseDokument52 SeitenDrugs For Parkinson's Diseasebhagyashri chaudhariNoch keine Bewertungen
- Etiology and Pathophysiology of Parkinson S DiseaseDokument552 SeitenEtiology and Pathophysiology of Parkinson S DiseaseAstrid Figueroa100% (2)
- Anti-Parkinson's Screening ModelsDokument64 SeitenAnti-Parkinson's Screening Modelsteenakunwar93% (15)
- Materi Sertifikasi TI UBDokument138 SeitenMateri Sertifikasi TI UBMaika100% (2)
- A Guide For The Assessment and Management of Post-Obstructive DiuresisDokument3 SeitenA Guide For The Assessment and Management of Post-Obstructive DiuresisRara Aulia IINoch keine Bewertungen
- Postobstructive Diuresis: Clinical ReviewDokument6 SeitenPostobstructive Diuresis: Clinical ReviewRara Aulia IINoch keine Bewertungen
- Post-Obstructive Diuresis: A Complication of Urinary RetentionDokument3 SeitenPost-Obstructive Diuresis: A Complication of Urinary RetentionRara Aulia IINoch keine Bewertungen
- E020500 FullDokument11 SeitenE020500 FullRara Aulia IINoch keine Bewertungen
- Bladder Decompression For Chronic Urinary Retention May Lead To Upper Tract BleedingDokument3 SeitenBladder Decompression For Chronic Urinary Retention May Lead To Upper Tract BleedingRara Aulia IINoch keine Bewertungen
- Pathophysiology and Management of Urinary Retention in Men: Review ArticleDokument7 SeitenPathophysiology and Management of Urinary Retention in Men: Review ArticleRara Aulia IINoch keine Bewertungen
- UC Irvine: Clinical Practice and Cases in Emergency MedicineDokument4 SeitenUC Irvine: Clinical Practice and Cases in Emergency MedicineRara Aulia IINoch keine Bewertungen
- Urinary Diversion Highlights: Editorial - 50th AnniversaryDokument12 SeitenUrinary Diversion Highlights: Editorial - 50th AnniversaryRara Aulia IINoch keine Bewertungen
- Full Text 01Dokument71 SeitenFull Text 01Rara Aulia IINoch keine Bewertungen
- Gynecologic Oncology: Omari K. Johnson, Andrew Berchuck, Angeles A. Secord, Andrew C. PetersonDokument8 SeitenGynecologic Oncology: Omari K. Johnson, Andrew Berchuck, Angeles A. Secord, Andrew C. PetersonRara Aulia IINoch keine Bewertungen
- Urinary Diversion After Pelvic Exenteration For Gynecologic MalignanciesDokument10 SeitenUrinary Diversion After Pelvic Exenteration For Gynecologic MalignanciesRara Aulia IINoch keine Bewertungen
- Dong 2015Dokument4 SeitenDong 2015Rara Aulia IINoch keine Bewertungen
- Ileal Neobladder and Its Variants: Georgios Gakis, Arnulf StenzlDokument9 SeitenIleal Neobladder and Its Variants: Georgios Gakis, Arnulf StenzlRara Aulia IINoch keine Bewertungen
- Lobo 2017Dokument10 SeitenLobo 2017Rara Aulia IINoch keine Bewertungen
- Robotic Urinary Diversion: The Range of Options: ReviewDokument7 SeitenRobotic Urinary Diversion: The Range of Options: ReviewRara Aulia IINoch keine Bewertungen
- UpdatesDokument24 SeitenUpdatesRara Aulia IINoch keine Bewertungen
- Daneshmand, 2017Dokument188 SeitenDaneshmand, 2017Rara Aulia IINoch keine Bewertungen
- New and Evolving Immunotherapy in Inflammatory Bowel DiseaseDokument11 SeitenNew and Evolving Immunotherapy in Inflammatory Bowel DiseaseRara Aulia IINoch keine Bewertungen
- Bibi 2017Dokument8 SeitenBibi 2017Rara Aulia IINoch keine Bewertungen
- Which Knife Was Used Using A Porcine Model To.2Dokument4 SeitenWhich Knife Was Used Using A Porcine Model To.2Rara Aulia IINoch keine Bewertungen
- A6 - B Ringkasan Kajian GiziDokument7 SeitenA6 - B Ringkasan Kajian GiziRara Aulia IINoch keine Bewertungen
- What Are The Advantages If We Culture Cell As Our Research Model and What Are The LimitationsDokument1 SeiteWhat Are The Advantages If We Culture Cell As Our Research Model and What Are The LimitationsRara Aulia IINoch keine Bewertungen
- Global Nutrition Report 2016Dokument180 SeitenGlobal Nutrition Report 2016IFPRINoch keine Bewertungen
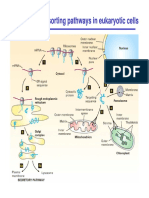
- Major Protein Sorting Pathways in Eukaryotic CellsDokument19 SeitenMajor Protein Sorting Pathways in Eukaryotic CellsRara Aulia IINoch keine Bewertungen
- Vitamin D AdministrationDokument1 SeiteVitamin D AdministrationRara Aulia IINoch keine Bewertungen
- Ijmcmed v3n4p216 enDokument9 SeitenIjmcmed v3n4p216 enRara Aulia IINoch keine Bewertungen
- HperlipidemiaDokument52 SeitenHperlipidemiaRara Aulia IINoch keine Bewertungen
- Rna ExtractionDokument21 SeitenRna ExtractionBaoz PingNoch keine Bewertungen
- Pediatric Clinics Type 1 Diabetes 2005Dokument26 SeitenPediatric Clinics Type 1 Diabetes 2005Rara Aulia IINoch keine Bewertungen
- Pharmacology of AutacoidsDokument13 SeitenPharmacology of AutacoidsInocenteNoch keine Bewertungen
- Dopaminergic Neurotoxicity of 1-Methyl-4-Phenyl-1,2,5,6 - Tetrahydropyridine in MiceDokument3 SeitenDopaminergic Neurotoxicity of 1-Methyl-4-Phenyl-1,2,5,6 - Tetrahydropyridine in MiceHamilton MorrisNoch keine Bewertungen
- Pathways of Parkinson's Disease from Pathology to MechanismsDokument13 SeitenPathways of Parkinson's Disease from Pathology to Mechanismsjust for download matterNoch keine Bewertungen
- 9497Dokument45 Seiten9497David axtonNoch keine Bewertungen
- Patogenesis ParkinsonDokument10 SeitenPatogenesis ParkinsonEric Yesaya TanNoch keine Bewertungen
- Expermental Models For Memory ImpairmentDokument13 SeitenExpermental Models For Memory ImpairmentIJAR JOURNALNoch keine Bewertungen
- Boldine and Its Antioxidant or Health-Promoting PropertiesDokument17 SeitenBoldine and Its Antioxidant or Health-Promoting PropertiesSleepyFishNoch keine Bewertungen
- Fearnley Lees BRAIN 1991Dokument19 SeitenFearnley Lees BRAIN 1991Sam StuartNoch keine Bewertungen
- Anti Park in Son MODELSDokument23 SeitenAnti Park in Son MODELSSameer KhanNoch keine Bewertungen
- Article WJPR 1538203814 PDFDokument13 SeitenArticle WJPR 1538203814 PDFAnant SrivastavaNoch keine Bewertungen
- Aerobic Exercise Enhances Neurodegenerative Diseases.Dokument2 SeitenAerobic Exercise Enhances Neurodegenerative Diseases.BRIGHT KWAKU ANYOMINoch keine Bewertungen
- S100B Protein Linked to Parkinson's Disease ProgressionDokument12 SeitenS100B Protein Linked to Parkinson's Disease ProgressionHasse HasseNoch keine Bewertungen
- 2019 - Rol de La MITOCONDRIA Y PARKINSONDokument92 Seiten2019 - Rol de La MITOCONDRIA Y PARKINSONEduardo RodriguezNoch keine Bewertungen
- Costa Et Al. - 2005 - Current Protocols in Toxicology - Wiley Interscience Eds PDFDokument2.760 SeitenCosta Et Al. - 2005 - Current Protocols in Toxicology - Wiley Interscience Eds PDFNicolas CancioNoch keine Bewertungen
- Combination of Honey and L-Dopa Protected The Dopaminergic Neurons Against MPTP Induced Parkinsonism in Adult Male Swiss MiceDokument6 SeitenCombination of Honey and L-Dopa Protected The Dopaminergic Neurons Against MPTP Induced Parkinsonism in Adult Male Swiss MiceInternational Journal of Innovative Science and Research Technology100% (1)
- Cellular and Molecular Mechanisms of Curcumin in Prevention and Treatment of DiseaseDokument55 SeitenCellular and Molecular Mechanisms of Curcumin in Prevention and Treatment of DiseaseJorge Luis Plasencia CubaNoch keine Bewertungen
- Zigmond MJ, Burke RE - Pathophysiology of Parkinson's DiseaseDokument14 SeitenZigmond MJ, Burke RE - Pathophysiology of Parkinson's Diseaseursula_ursula100% (1)
- Food and Nutrients in Disease Management - DepressionDokument21 SeitenFood and Nutrients in Disease Management - DepressionJJJPARKERNoch keine Bewertungen
- Parkinsonism (巴金森氏症) and other movement disordersDokument29 SeitenParkinsonism (巴金森氏症) and other movement disordersShing Ming TangNoch keine Bewertungen
- Parkinson's DiseaseDokument13 SeitenParkinson's DiseaseStella WatkinsNoch keine Bewertungen
- Halliwell 1992Dokument15 SeitenHalliwell 1992Prince MinatoNoch keine Bewertungen
- Resveratrol An Antiaging Drug With Potential Therapeutic Applications in Treating DiseasesDokument12 SeitenResveratrol An Antiaging Drug With Potential Therapeutic Applications in Treating DiseasesRenos DocNoch keine Bewertungen
- 2357 - KERUSAKAN OTAK DAN NEUROPLASTISITAS Biopsikologi 2018Dokument67 Seiten2357 - KERUSAKAN OTAK DAN NEUROPLASTISITAS Biopsikologi 2018Nurul HikmahNoch keine Bewertungen
- Magyar The Pharmacology of SelegilineDokument20 SeitenMagyar The Pharmacology of SelegilineAs DvgNoch keine Bewertungen
- Parkinsonism A General Motor Disability PDFDokument9 SeitenParkinsonism A General Motor Disability PDFRishabh SinghNoch keine Bewertungen