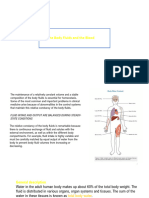
Das könnte Ihnen auch gefallen
- Renal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledVon EverandRenal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledNoch keine Bewertungen
- Cell-Associated Water: Proceedings of a Workshop on Cell-Associated Water Held in Boston, Massachusetts, September, 1976Von EverandCell-Associated Water: Proceedings of a Workshop on Cell-Associated Water Held in Boston, Massachusetts, September, 1976W. Drost-HansenNoch keine Bewertungen
- Water Physiology Essentiality, Metabolism, And.8Dokument6 SeitenWater Physiology Essentiality, Metabolism, And.8Patricia ChirinosNoch keine Bewertungen
- Darrow1950 PDFDokument9 SeitenDarrow1950 PDFLuís Eduardo CruvinelNoch keine Bewertungen
- Lectura N1A - The Discovery of Water Channels - AquaporinsDokument6 SeitenLectura N1A - The Discovery of Water Channels - AquaporinsJuanaNoch keine Bewertungen
- Emergency Fluid TherapyDokument7 SeitenEmergency Fluid TherapyAnaNoch keine Bewertungen
- Fluid, Electrolytes, Acid-Base and Shock ObjectivesDokument18 SeitenFluid, Electrolytes, Acid-Base and Shock ObjectivesmajedNoch keine Bewertungen
- Lab Activty On OsmoregulationDokument6 SeitenLab Activty On OsmoregulationKenneth Myro GarciaNoch keine Bewertungen
- Symposium: Proceedings I957Dokument5 SeitenSymposium: Proceedings I957Rachelle Mae Sonza AzarconNoch keine Bewertungen
- Fluid Electrolytes and Nutrition Physiological and Clinical AspectsDokument14 SeitenFluid Electrolytes and Nutrition Physiological and Clinical AspectsPaulynhiita FashyOnNoch keine Bewertungen
- Disorders of Water and Sodium BalanceDokument21 SeitenDisorders of Water and Sodium BalanceStanNoch keine Bewertungen
- Fluid and Electrolyte DisordersDokument9 SeitenFluid and Electrolyte DisordersRehab weridaNoch keine Bewertungen
- Sir David Cuthbertson Medal Lecture - Fluid, Electrolytes and Nutrition: Physiological and Clinical AspectsDokument15 SeitenSir David Cuthbertson Medal Lecture - Fluid, Electrolytes and Nutrition: Physiological and Clinical AspectsThinh VinhNoch keine Bewertungen
- PhysiologyDokument84 SeitenPhysiologyWilliam BufNoch keine Bewertungen
- Piis0007091217540933 PDFDokument6 SeitenPiis0007091217540933 PDFAde RohmanaNoch keine Bewertungen
- Perioperative Intravenous Fluid Therapy For Adults: Rob Mac Sweeney, Rachel Alexandra Mckendry, Amit BediDokument8 SeitenPerioperative Intravenous Fluid Therapy For Adults: Rob Mac Sweeney, Rachel Alexandra Mckendry, Amit BediAnnisa Chaerani BurhanuddinNoch keine Bewertungen
- Epidemiology, Pathophysiology, and Management of Hyponatremic EncephalopathyDokument11 SeitenEpidemiology, Pathophysiology, and Management of Hyponatremic EncephalopathyAnonymous Us5v7C6QhNoch keine Bewertungen
- Fluids and ElectrolytesDokument9 SeitenFluids and ElectrolytesCoy EnNoch keine Bewertungen
- Fluid Therapy Options and Rational SelectionDokument13 SeitenFluid Therapy Options and Rational SelectionRoberto López MataNoch keine Bewertungen
- Cell Tonicity LABDokument4 SeitenCell Tonicity LABa jNoch keine Bewertungen
- Etiology and Management of EdemaDokument14 SeitenEtiology and Management of EdemaFavio Alvarez De BejarNoch keine Bewertungen
- Mehu107 - U1 - T09 - Crystalloids, Colloids, Blood Products and Blood SubstitutesDokument8 SeitenMehu107 - U1 - T09 - Crystalloids, Colloids, Blood Products and Blood SubstitutesDANIEL ESTEBAN RODRIGUEZ TEJADANoch keine Bewertungen
- Disorders of Water, Electrolyte, and Mineral Ion MetabolismDokument4 SeitenDisorders of Water, Electrolyte, and Mineral Ion MetabolismSNoch keine Bewertungen
- Fluid and Electrolyte Disorders: Dr. Chandra Kant Pandey Dr. R. B. SinghDokument8 SeitenFluid and Electrolyte Disorders: Dr. Chandra Kant Pandey Dr. R. B. SinghMok Chu ZhenNoch keine Bewertungen
- Hypertonic Stress in The Kidney: A Necessary Evil: Min Seong Kwon, Sun Woo Lim and H. Moo KwonDokument7 SeitenHypertonic Stress in The Kidney: A Necessary Evil: Min Seong Kwon, Sun Woo Lim and H. Moo Kwoncrina1980Noch keine Bewertungen
- Water As An Essential Nutrient The PhysiologyDokument10 SeitenWater As An Essential Nutrient The PhysiologyGianinaMihalacheCojocaruNoch keine Bewertungen
- The American Journal of Medicine: EditorialDokument5 SeitenThe American Journal of Medicine: Editorialabidah fitraNoch keine Bewertungen
- CNUR823 - Module 2Dokument26 SeitenCNUR823 - Module 2AumNoch keine Bewertungen
- 03a Cell TonicityDokument4 Seiten03a Cell TonicityMary Jewel0% (1)
- Electrolytes PeriopDokument22 SeitenElectrolytes PeriopVivek PatangeNoch keine Bewertungen
- B.9 Situation - Care of Client With Problems in Fluid & Electrolyte BalanceDokument9 SeitenB.9 Situation - Care of Client With Problems in Fluid & Electrolyte BalanceSOLEIL LOUISE LACSON MARBASNoch keine Bewertungen
- Liquidos y Electrolitos 2Dokument4 SeitenLiquidos y Electrolitos 2Jorge MBNoch keine Bewertungen
- Oxygen Is Fundamental To LifeDokument5 SeitenOxygen Is Fundamental To LifemangiriNoch keine Bewertungen
- Urinary System EditedDokument8 SeitenUrinary System EditedJanith VimukthiNoch keine Bewertungen
- HEALTH SCI 12 - ANAPHY - Chapter - 27Dokument11 SeitenHEALTH SCI 12 - ANAPHY - Chapter - 27Kyla Marie BadanaNoch keine Bewertungen
- Disorders of Water ImbalanceDokument22 SeitenDisorders of Water ImbalanceRLauncherNoch keine Bewertungen
- Articulo Ingles III Fluid BalanceDokument4 SeitenArticulo Ingles III Fluid Balancesteve inga pomaNoch keine Bewertungen
- Lecture 14. Water - Mineral Metabolism. Biochemistry of Kidneys and UrineDokument39 SeitenLecture 14. Water - Mineral Metabolism. Biochemistry of Kidneys and UrineВіталій Михайлович НечипорукNoch keine Bewertungen
- The Physiological Regulation of Thirst and Fluid Intake: ArticlesDokument6 SeitenThe Physiological Regulation of Thirst and Fluid Intake: ArticlesCony GSNoch keine Bewertungen
- Intravenous Fluid TherapyDokument10 SeitenIntravenous Fluid TherapyAnonymous ysrxggk21cNoch keine Bewertungen
- Water-pH-Electrolytes EDITED!!! PDFDokument109 SeitenWater-pH-Electrolytes EDITED!!! PDFKate Lynne CamonayanNoch keine Bewertungen
- Fluids and Electrolytes: Epidemiology/PathophysiologyDokument27 SeitenFluids and Electrolytes: Epidemiology/PathophysiologyManna YankiNoch keine Bewertungen
- DwsDokument27 SeitenDwsKunni MardhiyahNoch keine Bewertungen
- Intraoperative Fluids: How Much Is Too Much?: Editor's Key PointsDokument11 SeitenIntraoperative Fluids: How Much Is Too Much?: Editor's Key PointsRicardoNoch keine Bewertungen
- Structured Water: The History, Science and Form For LifeDokument21 SeitenStructured Water: The History, Science and Form For Lifeministerenqi100% (2)
- Intracellular Fluid: Fluid Shifts Occur When The Body's Fluids Move Between The Fluid Compartments. Physiologically, ThisDokument4 SeitenIntracellular Fluid: Fluid Shifts Occur When The Body's Fluids Move Between The Fluid Compartments. Physiologically, ThisbryanpecundoballegaNoch keine Bewertungen
- Cellular Volume HomeostasisDokument6 SeitenCellular Volume HomeostasisAlberto AbaroaNoch keine Bewertungen
- Br. J. Anaesth. 2012 Doherty Bja - Aes171Dokument11 SeitenBr. J. Anaesth. 2012 Doherty Bja - Aes171Ahmad SaifulNoch keine Bewertungen
- Fluid Management: Basic ScienceDokument6 SeitenFluid Management: Basic ScienceDjeddah Illarik NoriegaNoch keine Bewertungen
- Fluids and ElectrolytesDokument8 SeitenFluids and ElectrolytesDEWINoch keine Bewertungen
- Intravenous Fluids and Electrolytes: Total Body Fluid CompositionDokument16 SeitenIntravenous Fluids and Electrolytes: Total Body Fluid Compositionaulia kamal ansari panggabeanNoch keine Bewertungen
- Balanced Versus Unbalanced Salt Solutions What (Raisul)Dokument13 SeitenBalanced Versus Unbalanced Salt Solutions What (Raisul)irmaarmiyahNoch keine Bewertungen
- Organization and Composition of Body FluidsDokument7 SeitenOrganization and Composition of Body FluidsEVELYN ITZEL OCHOA VARGASNoch keine Bewertungen
- Pediatric Fluid and Electrolyte Balance: Critical Care Case StudiesDokument13 SeitenPediatric Fluid and Electrolyte Balance: Critical Care Case StudiesSyarifa ZahraNoch keine Bewertungen
- Body Fluids and BloodDokument83 SeitenBody Fluids and Bloodalthea jade villadongaNoch keine Bewertungen
- 11.19 Excretory Products and Their ElimiiinationDokument6 Seiten11.19 Excretory Products and Their ElimiiinationMoonesh MKNoch keine Bewertungen
- F and e Imb .... Edu ..Dokument20 SeitenF and e Imb .... Edu ..esakkiammalNoch keine Bewertungen
- The Biological Importance of WaterDokument2 SeitenThe Biological Importance of WaterSophie Talib0% (3)
- Assessment of Fluid and ElectrolyteDokument15 SeitenAssessment of Fluid and ElectrolyteDip Ayan MNoch keine Bewertungen
- Honors Biology Critical Thinking Questions and Answers Chapter 5Dokument4 SeitenHonors Biology Critical Thinking Questions and Answers Chapter 5Henry Lawson80% (5)
- Rigel Uni Sim Lite DatasheetDokument5 SeitenRigel Uni Sim Lite Datasheetmailalexmd.vrn.ruNoch keine Bewertungen
- Asuhan Keperawatan (Severe Acute Respiratory Syndrome) : Ns. Kristina., MSNDokument25 SeitenAsuhan Keperawatan (Severe Acute Respiratory Syndrome) : Ns. Kristina., MSNMuhammad Sofyan NurNoch keine Bewertungen
- Organ Dysfunction in SepsisDokument19 SeitenOrgan Dysfunction in SepsisRossalia Yuliana100% (1)
- Overtraining Syndrome: A Practical Guide: Sports Health A Multidisciplinary Approach March 2012Dokument36 SeitenOvertraining Syndrome: A Practical Guide: Sports Health A Multidisciplinary Approach March 2012soyagurtNoch keine Bewertungen
- Class 4Dokument2 SeitenClass 4JackieNoch keine Bewertungen
- 1.case Presentation CHDDokument23 Seiten1.case Presentation CHDDHARM MEENANoch keine Bewertungen
- Bullworker and Elevated Blood PressureDokument3 SeitenBullworker and Elevated Blood PressureVenkitachalam RenganathanNoch keine Bewertungen
- Core CH 16 Coordination in Humans: 16.1.1 General Plan of The Nervous SystemDokument8 SeitenCore CH 16 Coordination in Humans: 16.1.1 General Plan of The Nervous SystemTSZ YAN CHEUNGNoch keine Bewertungen
- Jacob Gutierrez - Virtual Bioman PhotosynthesisDokument3 SeitenJacob Gutierrez - Virtual Bioman PhotosynthesisJacob GutierrezNoch keine Bewertungen
- High Acuity Exam 1 Study Guide - 2Dokument12 SeitenHigh Acuity Exam 1 Study Guide - 2Angelica WilliamsNoch keine Bewertungen
- Brain Wash: David Perlmutter, MD, and Austin Perlmutter, MDDokument7 SeitenBrain Wash: David Perlmutter, MD, and Austin Perlmutter, MDhanhdoc0% (1)
- CHAPTER 1 and 2 FINALDokument35 SeitenCHAPTER 1 and 2 FINALPrince Mark BadilloNoch keine Bewertungen
- Chapter 1: Respiratory and Circulatory System Working Together!Dokument5 SeitenChapter 1: Respiratory and Circulatory System Working Together!Precious Aiverose EspinaNoch keine Bewertungen
- Ion ChannelDokument15 SeitenIon ChannelWynona AlvarezNoch keine Bewertungen
- Pediatric Surgery Report 091118 FinalDokument33 SeitenPediatric Surgery Report 091118 FinalYuanico LiraukaNoch keine Bewertungen
- ACLS AHA Guidelines 2006 PosterDokument1 SeiteACLS AHA Guidelines 2006 PosterJeane SuyantoNoch keine Bewertungen
- Nutrition, Metabolism, and Body Temperature RegulationDokument11 SeitenNutrition, Metabolism, and Body Temperature RegulationSNoch keine Bewertungen
- X Bio Ch1 Life Processes Chapter NotesDokument20 SeitenX Bio Ch1 Life Processes Chapter NotesSurya Ravi0% (1)
- Acut Coronary SyndromeDokument65 SeitenAcut Coronary SyndromeArys SetiawanNoch keine Bewertungen
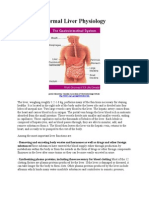
- Normal Liver PhysiologyDokument3 SeitenNormal Liver PhysiologymarinelleeslabonNoch keine Bewertungen
- Focus-Nursing Care Plan For Sleep DisordersDokument9 SeitenFocus-Nursing Care Plan For Sleep DisordersMae AzoresNoch keine Bewertungen
- Summative Test No. 2 ScienceDokument3 SeitenSummative Test No. 2 ScienceMelissa Favila PanagaNoch keine Bewertungen
- 9 Wireframe DocumentDokument7 Seiten9 Wireframe Documentharshaddanawale.nbnssoe.compNoch keine Bewertungen
- Excretion and OsmoregulationDokument48 SeitenExcretion and OsmoregulationMiguel Roman Reloaded Kerr100% (1)
- Hema 2 - Prelim Topic 2 - Additional Notes Primary HemostasisDokument2 SeitenHema 2 - Prelim Topic 2 - Additional Notes Primary HemostasisLowenstein JenzenNoch keine Bewertungen
- Nursing Care Plan For "Heart Failure Chronic"Dokument16 SeitenNursing Care Plan For "Heart Failure Chronic"jhonroks97% (34)
- Forensic 5 Group 4 ReportDokument13 SeitenForensic 5 Group 4 Report09476247345Noch keine Bewertungen
- G1 - Final Case Study - Anaphylaxis - BSN 3106Dokument43 SeitenG1 - Final Case Study - Anaphylaxis - BSN 3106Dj Gwyn MandigmaNoch keine Bewertungen
- Low Blood PressureDokument2 SeitenLow Blood PressurebambangNoch keine Bewertungen
- CPR and AED Final ExamDokument3 SeitenCPR and AED Final ExamNathan WhiteNoch keine Bewertungen