Das könnte Ihnen auch gefallen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Food Chemistry: Li-Chen Wu, Hsiu-Wen Hsu, Yun-Chen Chen, Chih-Chung Chiu, Yu-In Lin, Ja-An Annie HoDokument9 SeitenFood Chemistry: Li-Chen Wu, Hsiu-Wen Hsu, Yun-Chen Chen, Chih-Chung Chiu, Yu-In Lin, Ja-An Annie HoJanuarErlanggaNoch keine Bewertungen
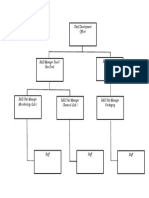
- Diagram StrukturDokument1 SeiteDiagram StrukturJanuarErlanggaNoch keine Bewertungen
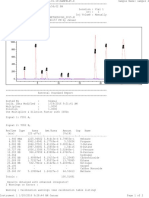
- Report Sample Dd.00sc5Dokument2 SeitenReport Sample Dd.00sc5JanuarErlanggaNoch keine Bewertungen
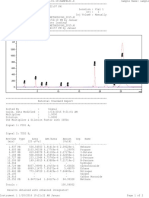
- Report Sample Aa.001Dokument2 SeitenReport Sample Aa.001JanuarErlanggaNoch keine Bewertungen
- Report Sample Aa.001Dokument2 SeitenReport Sample Aa.001JanuarErlanggaNoch keine Bewertungen
- Perlakuan Penurunan Kandungan Sianida Ubi Kayu Untuk Pakan TernakDokument4 SeitenPerlakuan Penurunan Kandungan Sianida Ubi Kayu Untuk Pakan TernakJanuarErlanggaNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Respiration in Plants Class 11 QuestionsDokument3 SeitenRespiration in Plants Class 11 QuestionsAshok KumarNoch keine Bewertungen
- Summary of PathwaysDokument1 SeiteSummary of Pathwaysayan100% (1)
- Chapter 19 - Gluconeogenesis & Amp The Control of Blood GlucoseDokument14 SeitenChapter 19 - Gluconeogenesis & Amp The Control of Blood Glucoserenzo041199Noch keine Bewertungen
- Development of A Gas-Phase...Dokument13 SeitenDevelopment of A Gas-Phase...otto11braunNoch keine Bewertungen
- Be209 Lab 3Dokument10 SeitenBe209 Lab 3Brian SoanNoch keine Bewertungen
- Metabolisme Karbohidrat.2020Dokument103 SeitenMetabolisme Karbohidrat.2020yayat yayatNoch keine Bewertungen
- Anaerobic Glycolysis in RBCDokument3 SeitenAnaerobic Glycolysis in RBCKanchana DissanayakeNoch keine Bewertungen
- Biochemistry 1.4 Enzymes Classification and KineticsDokument11 SeitenBiochemistry 1.4 Enzymes Classification and Kineticslovelots1234100% (1)
- Biology Worksheet For Remedial StudentsDokument25 SeitenBiology Worksheet For Remedial Studentsbaretoodo785Noch keine Bewertungen
- Worksheet: Cellular Respiration & Cell EnergyDokument4 SeitenWorksheet: Cellular Respiration & Cell Energyclaryl alexaNoch keine Bewertungen
- Gen Physics 1 2nd Quarter ExamDokument5 SeitenGen Physics 1 2nd Quarter ExamArvinNoch keine Bewertungen
- BW 1Dokument8 SeitenBW 1ChristinaNoch keine Bewertungen
- General BIology 1 Q2 M4 SCDokument15 SeitenGeneral BIology 1 Q2 M4 SCAldrin James DafunNoch keine Bewertungen
- Name: Decemei M. Cuabo. Year & Section: Bs-Biology 1St Yr. 1ADokument3 SeitenName: Decemei M. Cuabo. Year & Section: Bs-Biology 1St Yr. 1ADecemei CuaboNoch keine Bewertungen
- Biota Quiz Bee Mock Test Part 2Dokument2 SeitenBiota Quiz Bee Mock Test Part 2Ken Juliana Fe IsaacNoch keine Bewertungen
- Chapter 04Dokument11 SeitenChapter 04Wajeeh Ahmed ZakaiNoch keine Bewertungen
- WHLP Gen Bio 5 6 2nd QuarterDokument7 SeitenWHLP Gen Bio 5 6 2nd QuarterSir JoshNoch keine Bewertungen
- Cellular Respiration 9-12Dokument56 SeitenCellular Respiration 9-12David BayaniNoch keine Bewertungen
- HMP Pathway and Its Significance - PPTX Version 1Dokument40 SeitenHMP Pathway and Its Significance - PPTX Version 1Alana GeorgeNoch keine Bewertungen
- A Definition and Systems View of Anaerobic CapacityDokument7 SeitenA Definition and Systems View of Anaerobic CapacityRAMAMANI SAHOONoch keine Bewertungen
- Biochem Jsmu Ospe-1Dokument63 SeitenBiochem Jsmu Ospe-1Saif Ali100% (1)
- Bio Placement Test Review Questions Review 1: Answer Key On Page 11 Select The Correct AnswerDokument31 SeitenBio Placement Test Review Questions Review 1: Answer Key On Page 11 Select The Correct AnswerAndrea RaquilNoch keine Bewertungen
- Biology The Unity and Diversity of Life 12Th Edition Starr Test Bank Full Chapter PDFDokument33 SeitenBiology The Unity and Diversity of Life 12Th Edition Starr Test Bank Full Chapter PDFBeckySmithnxro100% (13)
- Energy and Respiration MSDokument52 SeitenEnergy and Respiration MSWaheed SherazNoch keine Bewertungen
- Dbmci CLTDokument284 SeitenDbmci CLTA. RaufNoch keine Bewertungen
- IB Unit PlanDokument11 SeitenIB Unit PlanChristina Dimitrijevic100% (2)
- Cell Energy SEDokument6 SeitenCell Energy SEArt LoversNoch keine Bewertungen
- GlycolysisDokument21 SeitenGlycolysisAli AlqumaNoch keine Bewertungen
- CC - DAY 2 - PRE-TEST RationalizationDokument30 SeitenCC - DAY 2 - PRE-TEST RationalizationVincent AmitNoch keine Bewertungen
- Chapter 05Dokument80 SeitenChapter 05John Phillip DionisioNoch keine Bewertungen