Das könnte Ihnen auch gefallen
- Manual de Programación OKUMA EspañolDokument382 SeitenManual de Programación OKUMA Españolmaiden80% (5)
- Taller On LineDokument4 SeitenTaller On Lineelijanet50% (2)
- Analisis de La OclusionDokument5 SeitenAnalisis de La OclusionCarla Silvana Alvarado CostaNoch keine Bewertungen
- Evaluacion 1 Analisis SensorialDokument3 SeitenEvaluacion 1 Analisis SensorialDenis Huanca De La CruzNoch keine Bewertungen
- Micro 3Dokument10 SeitenMicro 3AlexNoch keine Bewertungen
- Lipotimia y Sincope 1Dokument25 SeitenLipotimia y Sincope 1Luis Aarón Cabello CandelaNoch keine Bewertungen
- Medicamentos AntipireticosDokument6 SeitenMedicamentos AntipireticosPaola Huamán FloresNoch keine Bewertungen
- Reconocimiento de Sabores BasicoDokument8 SeitenReconocimiento de Sabores BasicoIsaac DavidNoch keine Bewertungen
- Mecanismos Neuromusculares de Control de La Función MasticatoriaDokument2 SeitenMecanismos Neuromusculares de Control de La Función MasticatoriamarojassNoch keine Bewertungen
- Fisiología Del HuesoDokument7 SeitenFisiología Del HuesoKAREN DANIELA PARADA PALACIOS100% (2)
- Informe - ProteinasDokument8 SeitenInforme - ProteinasCintia Azucena Villanueva EstacioNoch keine Bewertungen
- Ácidos Orgánicos Producidos Por FermentaciónDokument27 SeitenÁcidos Orgánicos Producidos Por FermentaciónDerly Loaiza Ninantay100% (1)
- Conceptos Basicos de MicrobiologíaDokument11 SeitenConceptos Basicos de MicrobiologíaReinaldo Morales ForeroNoch keine Bewertungen
- Glucolisis 211Dokument38 SeitenGlucolisis 211Fernando Tadeo CapistranNoch keine Bewertungen
- ArticuladoresDokument17 SeitenArticuladoresRoriana RangelNoch keine Bewertungen
- Agentes Físicos para El Control de MicroorganismosDokument9 SeitenAgentes Físicos para El Control de MicroorganismosCarla Noriega PerezNoch keine Bewertungen
- Etapas Del Anabolismo y CatabolismoDokument8 SeitenEtapas Del Anabolismo y CatabolismoAxel MelchorNoch keine Bewertungen
- Re Tenedor EsDokument37 SeitenRe Tenedor EsAndres OrtizNoch keine Bewertungen
- Ortesis CervicalesDokument6 SeitenOrtesis CervicalesArturo Bazan100% (1)
- Normas de Bioseguridad InformeDokument4 SeitenNormas de Bioseguridad Informemaried eliana marin50% (2)
- Atrición DentalDokument3 SeitenAtrición DentalMaría Cuñas RamosNoch keine Bewertungen
- RECURSODokument1 SeiteRECURSOmarcosNoch keine Bewertungen
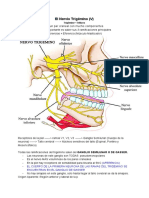
- Par Craneal 5 PDFDokument8 SeitenPar Craneal 5 PDFAlicia Trimmiño JaramilloNoch keine Bewertungen
- Seminario de Bioetica Valor y Antivalor - Semestre 1 - 2019Dokument18 SeitenSeminario de Bioetica Valor y Antivalor - Semestre 1 - 2019Lina Maria Malo GomezNoch keine Bewertungen
- Ensayo de Operatoria DentalDokument3 SeitenEnsayo de Operatoria Dentaljonathan100% (1)
- Evaluacion SensorialDokument64 SeitenEvaluacion SensorialRosy TopeteNoch keine Bewertungen
- Alteraciones en La Cinemática MandibularDokument6 SeitenAlteraciones en La Cinemática MandibularanabelNoch keine Bewertungen
- Semana 3 - Material de Trabajo (202120)Dokument4 SeitenSemana 3 - Material de Trabajo (202120)Diego WongNoch keine Bewertungen
- Cubetas de Impresión Finales - WebDokument12 SeitenCubetas de Impresión Finales - WebIrsa Liz100% (1)
- Otitis Externa y MediaDokument6 SeitenOtitis Externa y MediaCarlos RubioNoch keine Bewertungen
- Actividad 1. Mapa Conceptual.Dokument3 SeitenActividad 1. Mapa Conceptual.Pamela JeronimoNoch keine Bewertungen
- ENVEJECIMIENTO - Cambios Bioquímicos - ZentenoDokument7 SeitenENVEJECIMIENTO - Cambios Bioquímicos - ZentenoCarlos Albert Krus100% (1)
- Indices Placa BacterianaDokument14 SeitenIndices Placa BacterianaMONICA DANIELA ROMERO NOCUANoch keine Bewertungen
- AmoxibrolDokument17 SeitenAmoxibrolkarinaNoch keine Bewertungen
- AntimicrobianosDokument9 SeitenAntimicrobianosLenin Franco TorayaNoch keine Bewertungen
- Equipos de LaboratorioDokument24 SeitenEquipos de Laboratoriofernando abraham berrospi casano100% (1)
- Practica de Embriologia DentariaDokument12 SeitenPractica de Embriologia DentariaGerardo Bardales ArgüellesNoch keine Bewertungen
- FitohemaglutininaDokument9 SeitenFitohemaglutininaChristina ZtaNoch keine Bewertungen
- Clase 3Dokument13 SeitenClase 3Leonardo Jesus Torres OjedaNoch keine Bewertungen
- Eden Tuli SmoDokument32 SeitenEden Tuli SmoMary Lisset Bermeo FloresNoch keine Bewertungen
- Farm Gastro IntDokument68 SeitenFarm Gastro Intsesa123Noch keine Bewertungen
- Mapa Conceptual Habitos Orales, HC 1.Dokument1 SeiteMapa Conceptual Habitos Orales, HC 1.Diana Rodríguez PuertaNoch keine Bewertungen
- Organos Linfoides 2016 1Dokument19 SeitenOrganos Linfoides 2016 1Diego Iván De La CruzNoch keine Bewertungen
- LAMINA PROPIA - HistologiaDokument2 SeitenLAMINA PROPIA - HistologiaKarina ValdiglesiasNoch keine Bewertungen
- Clase IV Morfofisiologia de La Cavidad OralDokument6 SeitenClase IV Morfofisiologia de La Cavidad OralIngrid Marcela León AlvarezNoch keine Bewertungen
- Nervio EspinalDokument6 SeitenNervio EspinalSabrina DiazNoch keine Bewertungen
- Dolor de Cuello o Cervicalgia - FisiotecaDokument3 SeitenDolor de Cuello o Cervicalgia - FisiotecaMartin EllinghamNoch keine Bewertungen
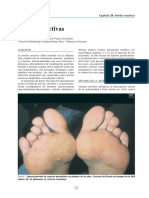
- Cap 28 Artritis Reactivas PDFDokument12 SeitenCap 28 Artritis Reactivas PDFNatii CampOsNoch keine Bewertungen
- Vademecum Medical Nutrition PDFDokument214 SeitenVademecum Medical Nutrition PDFMaricruz Saldaña100% (1)
- Alveogyl Ficha Resolución PDFDokument1 SeiteAlveogyl Ficha Resolución PDFGaia VarezNoch keine Bewertungen
- Corticoesteroides o Corticoides ExpoDokument3 SeitenCorticoesteroides o Corticoides ExpoTatiana SanchezNoch keine Bewertungen
- 16 QuerionDokument3 Seiten16 QuerionAxoncomunicacion100% (1)
- Presentación 1Dokument13 SeitenPresentación 1Luis VargasNoch keine Bewertungen
- Fisuras Segun KernahanDokument18 SeitenFisuras Segun KernahanGonzalo Molina GonzalezNoch keine Bewertungen
- Trastornos Dentales Inducidos Por Farmacos PDFDokument4 SeitenTrastornos Dentales Inducidos Por Farmacos PDFBLINDED185231Noch keine Bewertungen
- Epigastralgias PDFDokument14 SeitenEpigastralgias PDFHugo Gutiérrez MoralesNoch keine Bewertungen
- Cavidad Oral InformeDokument8 SeitenCavidad Oral InformevabyavilaNoch keine Bewertungen
- Canino InferiorDokument1 SeiteCanino InferiorAnonymous iVQXw9PWNoch keine Bewertungen
- BLASTOMICOSISDokument15 SeitenBLASTOMICOSISCesar Alfredo Cobeñas VillarrealNoch keine Bewertungen
- HEMICELULOSADokument22 SeitenHEMICELULOSAAryanna Mormont0% (1)
- La Fisiologia de Los SentidosDokument18 SeitenLa Fisiologia de Los Sentidosrociocatalina56Noch keine Bewertungen
- Fisiología de La VisiónDokument6 SeitenFisiología de La VisiónMazuey CroissantsNoch keine Bewertungen
- LuuuuuuuuuDokument1 SeiteLuuuuuuuuuPaula Fernanda PrietoNoch keine Bewertungen
- Cinematica Del TraumaDokument3 SeitenCinematica Del TraumaelijanetNoch keine Bewertungen
- PRT CaidasDokument1 SeitePRT CaidaselijanetNoch keine Bewertungen
- Escala BrademDokument1 SeiteEscala BrademCarmenxuu Durán MarileoNoch keine Bewertungen
- Poema Noche SobreDokument1 SeitePoema Noche SobreelijanetNoch keine Bewertungen
- Oda A Una Estrella PoemaDokument1 SeiteOda A Una Estrella PoemaelijanetNoch keine Bewertungen
- EFAMchileDokument6 SeitenEFAMchileDaniel RebolledoNoch keine Bewertungen
- EFAMchileDokument6 SeitenEFAMchileDaniel RebolledoNoch keine Bewertungen
- Poema Lo Que El Dinero Puede ComprarDokument1 SeitePoema Lo Que El Dinero Puede ComprarelijanetNoch keine Bewertungen
- Medicamentos Farmacos Resumen 1Dokument5 SeitenMedicamentos Farmacos Resumen 1elijanetNoch keine Bewertungen
- Sistemas Estructurales 1Dokument54 SeitenSistemas Estructurales 1Dulce DCNoch keine Bewertungen
- La Relatividad PDFDokument10 SeitenLa Relatividad PDFSantiago CamposNoch keine Bewertungen
- Segundo Trabajo Integrador de Recuperación de Saberes de Segundo AñoDokument2 SeitenSegundo Trabajo Integrador de Recuperación de Saberes de Segundo AñoJulï CoriaNoch keine Bewertungen
- Trabajo de Hidrostatica F. IiDokument8 SeitenTrabajo de Hidrostatica F. IiJorge Luis Vásquez TeodoroNoch keine Bewertungen
- FORMULAS EMPIRICAS SEDIMENTOSpdfDokument78 SeitenFORMULAS EMPIRICAS SEDIMENTOSpdfRolando Murillo RoqueNoch keine Bewertungen
- Fasproblemas de AnalisisDokument21 SeitenFasproblemas de AnalisisGracePostigoTheranNoch keine Bewertungen
- Investigacion Sobre TransformadoresDokument21 SeitenInvestigacion Sobre TransformadoresWilliam LemNoch keine Bewertungen
- Tema 5 - Depuracià N Gases - 2019-20Dokument36 SeitenTema 5 - Depuracià N Gases - 2019-20ali cherradiNoch keine Bewertungen
- Cuestionario Previo p3 FMDokument6 SeitenCuestionario Previo p3 FMhavoNoch keine Bewertungen
- ELECTRONICA I (4ta)Dokument7 SeitenELECTRONICA I (4ta)Claudia CalderonNoch keine Bewertungen
- Cuestionario N°1-Resistencia de Materiales T-ADokument4 SeitenCuestionario N°1-Resistencia de Materiales T-AAlvaro RevillaNoch keine Bewertungen
- Ficha Tecnica Escalera Multiproposito Tipo IDokument1 SeiteFicha Tecnica Escalera Multiproposito Tipo IFRANCISCO100% (1)
- Carreño Montiel Sebastian Proyecto de DiseñoDokument37 SeitenCarreño Montiel Sebastian Proyecto de Diseñodanny mottaNoch keine Bewertungen
- U4 TermodinámicaDokument74 SeitenU4 TermodinámicaEddy Ipanaque PerezNoch keine Bewertungen
- Instalaciones Sanitarias Grupo BDokument8 SeitenInstalaciones Sanitarias Grupo BAh LiamNoch keine Bewertungen
- Foro Debate TermodinamicaDokument2 SeitenForo Debate TermodinamicaEdwin Vilchez SolanoNoch keine Bewertungen
- Procedimiento de Calidad Instalaciones Electricas 01Dokument15 SeitenProcedimiento de Calidad Instalaciones Electricas 01Fernando PajaresNoch keine Bewertungen
- Métodos de Síntesis de NanopartículasDokument14 SeitenMétodos de Síntesis de NanopartículasHeber Andrada86% (7)
- Ciclo Del AguaDokument6 SeitenCiclo Del AguaALARCON BANDA HERMESNoch keine Bewertungen
- Sáenz AzorsaDokument161 SeitenSáenz AzorsaFranz Limber MendozaNoch keine Bewertungen
- Elementos de Laboratorio Fisica III - Daniel Rico FonsecaDokument6 SeitenElementos de Laboratorio Fisica III - Daniel Rico FonsecaDANIEL ALEJANDRO RICO FONSECANoch keine Bewertungen
- WEG Micro y Mini Drives ManualDokument174 SeitenWEG Micro y Mini Drives ManualPabloNoch keine Bewertungen
- 05 ProyectilesDokument18 Seiten05 Proyectilesramirodecristo2Noch keine Bewertungen
- Análisis Energético de Redes de Aire ComprimidoDokument23 SeitenAnálisis Energético de Redes de Aire ComprimidojvasquezespNoch keine Bewertungen
- PRIM Libro ChirifDokument228 SeitenPRIM Libro ChirifjordanNoch keine Bewertungen
- RetrieveDokument79 SeitenRetrieveYesi NievasNoch keine Bewertungen
- Caida InclinadaDokument3 SeitenCaida InclinadaOswaldo De la CruzNoch keine Bewertungen
- Transformacion de Esfuerzos - ECU - TransformacionDokument4 SeitenTransformacion de Esfuerzos - ECU - TransformacionJordy EucedaNoch keine Bewertungen
- Expediente Instalaciones EléctricasDokument42 SeitenExpediente Instalaciones EléctricascesarNoch keine Bewertungen