Das könnte Ihnen auch gefallen
- Requisitos de Equivalencia PDFDokument1 SeiteRequisitos de Equivalencia PDFSam HidalgoNoch keine Bewertungen
- Recaudos Exigidos para SolicitudDokument1 SeiteRecaudos Exigidos para SolicitudSam HidalgoNoch keine Bewertungen
- Como Superar Una Infidelidad Con La Ayuda de La Pareja Que Fue InfielDokument27 SeitenComo Superar Una Infidelidad Con La Ayuda de La Pareja Que Fue Infielsentranic100% (2)
- Requisitos de EquivalenciaDokument1 SeiteRequisitos de EquivalenciaSam HidalgoNoch keine Bewertungen
- Curriculun VitaeDokument1 SeiteCurriculun VitaeMiqueas Malaquías Velásquez CondoríNoch keine Bewertungen
- Curriculun VitaeDokument1 SeiteCurriculun VitaeMiqueas Malaquías Velásquez CondoríNoch keine Bewertungen
- Tema 3 Bioenergetica, MedicinaDokument13 SeitenTema 3 Bioenergetica, MedicinaSam HidalgoNoch keine Bewertungen
- Nociones de Embriologia FecundacionDokument22 SeitenNociones de Embriologia FecundacionSam HidalgoNoch keine Bewertungen
- I Unidad Morfofisiologia III Completa Samuel ReyesDokument186 SeitenI Unidad Morfofisiologia III Completa Samuel ReyesSam Hidalgo90% (10)
- Pae 1Dokument5 SeitenPae 1adelafuenteNoch keine Bewertungen
- Cuello Anatomia DefinitivoDokument58 SeitenCuello Anatomia DefinitivoCristian Pino100% (2)
- AuriculoterapiaDokument46 SeitenAuriculoterapialago354Noch keine Bewertungen
- Sistema Digestivo PoligastricoDokument24 SeitenSistema Digestivo PoligastricoRonald Cardozo UrzagasteNoch keine Bewertungen
- Monografia Lactancia MaternaDokument34 SeitenMonografia Lactancia MaternaobedNoch keine Bewertungen
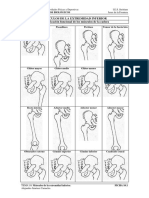
- Músculos de La Cadera PDFDokument2 SeitenMúsculos de La Cadera PDFMarcosPrietoVeizagaNoch keine Bewertungen
- (T4DW) Sword Art Online Volumen 9 Prólogo 2 (V-Normal) PDFDokument0 Seiten(T4DW) Sword Art Online Volumen 9 Prólogo 2 (V-Normal) PDFAlberto del RioNoch keine Bewertungen
- Taller de Fisiologia RespiratorioDokument16 SeitenTaller de Fisiologia RespiratorioJuan Esteban AYALA ROMERONoch keine Bewertungen
- U.F 119 PDFDokument105 SeitenU.F 119 PDFCami Barba LópezNoch keine Bewertungen
- Mapa Conceptual Del Sistema Respiratorio 2Dokument1 SeiteMapa Conceptual Del Sistema Respiratorio 2Abril LamasNoch keine Bewertungen
- Fisiologia y Componentes Del Sistema Circulatorio y Sus FuncionesDokument15 SeitenFisiologia y Componentes Del Sistema Circulatorio y Sus FuncionesMaria Gabriela Cuevas BastidasNoch keine Bewertungen
- Informe-2 MayteDokument8 SeitenInforme-2 MayteRodolfo LunaNoch keine Bewertungen
- Enfermedades Parasitarias de Las AvesDokument86 SeitenEnfermedades Parasitarias de Las AvesDavis Garcia67% (3)
- Cardiopatias Congenitas 2019Dokument101 SeitenCardiopatias Congenitas 2019ThaliaMirandaNoch keine Bewertungen
- EMBRIOGENESISDokument34 SeitenEMBRIOGENESISMarcelo Arancibia VidelaNoch keine Bewertungen
- Ivu NotaDokument1 SeiteIvu NotaJo Ker Puddin100% (1)
- Estructura y Funcion Del CuerpoDokument7 SeitenEstructura y Funcion Del CuerpoJassiel AlfaroNoch keine Bewertungen
- SIMCEDokument8 SeitenSIMCEAgustin Valdes SanchezNoch keine Bewertungen
- Clase 9 Fotosintesis de C3 y C4Dokument27 SeitenClase 9 Fotosintesis de C3 y C4GUEVARA GALVEZ LEIDYNoch keine Bewertungen
- 2430-005-002 Instrucciones para La Medici+ N de Signos Vitales y Determinaci+ N de Somatometr+¡aDokument19 Seiten2430-005-002 Instrucciones para La Medici+ N de Signos Vitales y Determinaci+ N de Somatometr+¡aJulio César Vázquez BustosNoch keine Bewertungen
- Conceptos de Higiene PecuariaDokument34 SeitenConceptos de Higiene PecuariaVianey Melendez60% (5)
- Indicaciones Especificas de HemocomponentesDokument39 SeitenIndicaciones Especificas de HemocomponentesAlvaro Danilño Leiva López100% (1)
- Glosario de EnfermeriaDokument25 SeitenGlosario de EnfermeriaJossj93100% (1)
- Botanica Practica I-2015 ImprimirDokument7 SeitenBotanica Practica I-2015 ImprimirJuan Luis Huertas MendozaNoch keine Bewertungen
- Biologia Listo 3Dokument4 SeitenBiologia Listo 3FABIÁN Ponce GómezNoch keine Bewertungen
- Prueba Ciencias Naturales Sistema LocomotorDokument4 SeitenPrueba Ciencias Naturales Sistema LocomotorJaviera Lorena Sanhueza CarreraNoch keine Bewertungen
- Masoterapia-Sistema NerviosoDokument7 SeitenMasoterapia-Sistema NerviosonoeliaNoch keine Bewertungen
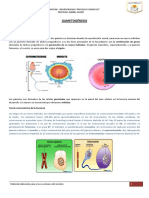
- Que Es La EndorfinaDokument1 SeiteQue Es La EndorfinaJuan Carlos Chinchilla MuñozNoch keine Bewertungen
- Análisis de Control de Calidad en Incubación de HuevosDokument31 SeitenAnálisis de Control de Calidad en Incubación de HuevosGuillermo Zavala ANoch keine Bewertungen
- Mapa ConceptualDokument1 SeiteMapa ConceptualGabriela RoaNoch keine Bewertungen