Das könnte Ihnen auch gefallen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Smythe1997 MR Efeito AnsioliticoDokument7 SeitenSmythe1997 MR Efeito AnsioliticoKairo AlbernazNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Lachman 1997Dokument6 SeitenLachman 1997Kairo AlbernazNoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Lachman 1997Dokument6 SeitenLachman 1997Kairo AlbernazNoch keine Bewertungen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- De Kloet 1996Dokument7 SeitenDe Kloet 1996Kairo AlbernazNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Temporal Analysis of The Rat's Behavior in The Plus-Maze: Effect of MidazolamDokument6 SeitenTemporal Analysis of The Rat's Behavior in The Plus-Maze: Effect of MidazolamKairo AlbernazNoch keine Bewertungen
- Baptista 2012Dokument6 SeitenBaptista 2012Kairo AlbernazNoch keine Bewertungen
- The History of Benzodiazepines: in PracticeDokument10 SeitenThe History of Benzodiazepines: in PracticeKairo AlbernazNoch keine Bewertungen
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- J EuroneuroDokument11 SeitenJ EuroneuroKairo AlbernazNoch keine Bewertungen
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The History of Benzodiazepines: in PracticeDokument10 SeitenThe History of Benzodiazepines: in PracticeKairo AlbernazNoch keine Bewertungen
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Antihyperalgesic Antinociceptive Effects Of.32 PDFDokument14 SeitenAntihyperalgesic Antinociceptive Effects Of.32 PDFKairo AlbernazNoch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Nesse R. 1998. Emotional Disorders in Evolutionary PerspectiveDokument19 SeitenNesse R. 1998. Emotional Disorders in Evolutionary PerspectiveKairo AlbernazNoch keine Bewertungen
- 5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)Dokument115 Seiten5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)MobarobberNoch keine Bewertungen
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Colegio de San Juan de LetranDokument71 SeitenColegio de San Juan de Letranirish xNoch keine Bewertungen
- SassafasDokument462 SeitenSassafashasla18Noch keine Bewertungen
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Definition of Parental CareDokument8 SeitenDefinition of Parental CareLemon GundersonNoch keine Bewertungen
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
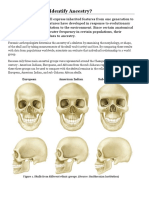
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanDokument5 SeitenActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Plant StructureDokument6 SeitenPlant StructureYoga Trista MajidNoch keine Bewertungen
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Pharmaceutical Microbiology - CCDokument3 SeitenPharmaceutical Microbiology - CCRahardi MokhtarNoch keine Bewertungen
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Cerebellum-8 PDFDokument83 SeitenThe Cerebellum-8 PDFlovelots1234Noch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Ch42 Practice TestDokument8 SeitenCh42 Practice TestMike Wolsky0% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Atf6 - 73505ex.20100921Dokument3 SeitenAtf6 - 73505ex.20100921Alyssa236Noch keine Bewertungen
- Euglena) Some Colonial (Volvox)Dokument22 SeitenEuglena) Some Colonial (Volvox)Faiqoh NahareyNoch keine Bewertungen
- Grzimek's Animal Life EncyclopediaDokument2 SeitenGrzimek's Animal Life Encyclopediamaximilijan7Noch keine Bewertungen
- Npre2007241 2 PDFDokument134 SeitenNpre2007241 2 PDFranjeet kumarNoch keine Bewertungen
- CLA Cattle Market Report September 29, 2021Dokument1 SeiteCLA Cattle Market Report September 29, 2021Clovis Livestock AuctionNoch keine Bewertungen
- Land AnimalsDokument52 SeitenLand AnimalsVinuta AilNoch keine Bewertungen
- đề thi preDokument14 Seitenđề thi prePham congNoch keine Bewertungen
- Animal Kingdom BiologyDokument8 SeitenAnimal Kingdom BiologyMary Ann SalvatierraNoch keine Bewertungen
- Head and Neck Anatomy 1Dokument7 SeitenHead and Neck Anatomy 1an leNoch keine Bewertungen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Reptiles & Amphibians: Second Report of Ophiophagy in A Cat-Eyed Snake (Leptodeira SP.) in Costa RicaDokument2 SeitenReptiles & Amphibians: Second Report of Ophiophagy in A Cat-Eyed Snake (Leptodeira SP.) in Costa Ricaapi-309280225Noch keine Bewertungen
- Student's Book: Kim MilneDokument17 SeitenStudent's Book: Kim MilneЕлизавета БогоявленскаяNoch keine Bewertungen
- 5 A. Floral Formulas and DiagramsDokument30 Seiten5 A. Floral Formulas and DiagramsGhanshyam R. Parmar33% (3)
- Kedah Matriculation College: BiodiversityDokument12 SeitenKedah Matriculation College: BiodiversityMaisarahNoch keine Bewertungen
- Chat 2Dokument1.730 SeitenChat 2Mujeeb LoopzNoch keine Bewertungen
- Membrane TransportDokument74 SeitenMembrane TransportMiftahul KhairaniNoch keine Bewertungen
- Animal KingdomDokument67 SeitenAnimal KingdomNaveen Krishna M.KNoch keine Bewertungen
- Three Essays On The Dinosaurs - Immanuel VelikovskyDokument6 SeitenThree Essays On The Dinosaurs - Immanuel VelikovskyBright StarrNoch keine Bewertungen
- Simon LeVay The Sexual Brain 1994Dokument156 SeitenSimon LeVay The Sexual Brain 1994acadlee100% (3)
- JKSSB Class IV 27 Feb Paper JK Latest InfoDokument12 SeitenJKSSB Class IV 27 Feb Paper JK Latest InfoIdrees AhmedNoch keine Bewertungen
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- English - (Uasa) f1 - English Model 1 - 2023Dokument12 SeitenEnglish - (Uasa) f1 - English Model 1 - 2023m-11267738Noch keine Bewertungen
- English 3RD TestDokument3 SeitenEnglish 3RD TestMichael CalesajrNoch keine Bewertungen