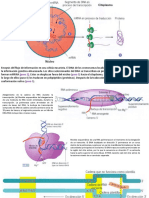
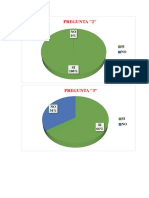
Das könnte Ihnen auch gefallen
- Practica # 1 Las MutacionesDokument8 SeitenPractica # 1 Las MutacionesRania ArbajeNoch keine Bewertungen
- Regulación Génica en EucariotesDokument44 SeitenRegulación Génica en EucariotesMauricio SanchezNoch keine Bewertungen
- Regulación de La Expresión Génica EucariotasDokument20 SeitenRegulación de La Expresión Génica EucariotasGloribel Bolívar100% (1)
- BIOLOGIA MOLECULAR Y NEFROLOGIA Análisis Del DNADokument19 SeitenBIOLOGIA MOLECULAR Y NEFROLOGIA Análisis Del DNASANTIAGO DENIS INTURIAS RODRIGUEZNoch keine Bewertungen
- Control Transcripcional de La Expresion GeneticaDokument2 SeitenControl Transcripcional de La Expresion Geneticaalexandra mazza100% (1)
- Los Estimuladores y Silenciadores Elementos de RegulaciónDokument3 SeitenLos Estimuladores y Silenciadores Elementos de RegulaciónDIEGO MAURICIO SALAZAR ZAMBRANONoch keine Bewertungen
- Transcripción y TraducciónDokument6 SeitenTranscripción y TraducciónJose Luis VallejosNoch keine Bewertungen
- Silo - Tips - La Transcripcion en EucariotasDokument25 SeitenSilo - Tips - La Transcripcion en EucariotasDai DinamarcaNoch keine Bewertungen
- Práctica N1 Partes de Un GenDokument9 SeitenPráctica N1 Partes de Un GenArian Alejandra Becerra RodriguezNoch keine Bewertungen
- Clase 13 Traducción - Regulación Expresión-BiolDokument53 SeitenClase 13 Traducción - Regulación Expresión-BiolKatherinne Perez SanchezNoch keine Bewertungen
- Parcial Genética 2020 C - RDokument14 SeitenParcial Genética 2020 C - RGuille DiazNoch keine Bewertungen
- Taller 6. Regulación de La Expresión en EucariotasDokument4 SeitenTaller 6. Regulación de La Expresión en EucariotasPaula NastarNoch keine Bewertungen
- La Regulación de La Traducción de ProteínasDokument4 SeitenLa Regulación de La Traducción de ProteínasLUISA DANIELA CACERES RUBIONoch keine Bewertungen
- Cap. 5 - TranscripcionDokument7 SeitenCap. 5 - TranscripcionJaqueline CavalcanteNoch keine Bewertungen
- Estructura Molecular Del Gen EucariotaDokument6 SeitenEstructura Molecular Del Gen Eucariotagvivana100% (1)
- BiologiaDokument11 SeitenBiologiaLaura SierraNoch keine Bewertungen
- Metabolismo de RNADokument40 SeitenMetabolismo de RNACesar Pariona100% (1)
- PlasmidosDokument23 SeitenPlasmidosBetiana BerriryNoch keine Bewertungen
- Taller Control de La Expresión GenicaDokument4 SeitenTaller Control de La Expresión Genicakike1600Noch keine Bewertungen
- Te AmoDokument6 SeitenTe AmoAliss LilibethNoch keine Bewertungen
- InsulinaDokument12 SeitenInsulinaErnesto ArguelloNoch keine Bewertungen
- Presentacion Geralk Naturaleza Del GenDokument28 SeitenPresentacion Geralk Naturaleza Del GenWaldo Guillermo RuizNoch keine Bewertungen
- Calvopina Jonatan Resumen ArticuloDokument2 SeitenCalvopina Jonatan Resumen ArticuloOvni MaxNoch keine Bewertungen
- BioquiDokument9 SeitenBioquiGiovanni GregorioNoch keine Bewertungen
- Taller. Regulación de La Expresión en Eucariotas1 (2) 1 1Dokument5 SeitenTaller. Regulación de La Expresión en Eucariotas1 (2) 1 1Santiago MartinezNoch keine Bewertungen
- POLYOMAVIRUSES 2 EsDokument11 SeitenPOLYOMAVIRUSES 2 EsMICHELL CAROLINA FERNANDEZ PAEZNoch keine Bewertungen
- Síntesis de Proteínas (Bioquímica 2009)Dokument56 SeitenSíntesis de Proteínas (Bioquímica 2009)RosarioJuyoSalazarNoch keine Bewertungen
- Transcripción en procariotas: regulación y mecanismosDokument5 SeitenTranscripción en procariotas: regulación y mecanismosSuárez Acevedo C. SaritaNoch keine Bewertungen
- TranscripciónDokument68 SeitenTranscripciónMehida Mishell Buch YaxónNoch keine Bewertungen
- Proceso de Transcripción y Su RegulacionDokument4 SeitenProceso de Transcripción y Su RegulacionKevin Andres Z.Noch keine Bewertungen
- Cómo operan los mecanismos epigenéticos en la regulación de la expresión génicaDokument6 SeitenCómo operan los mecanismos epigenéticos en la regulación de la expresión génicaJeniffer SalazarNoch keine Bewertungen
- Nutrigenómica y nutrigenética: la base genética de la nutriciónDokument53 SeitenNutrigenómica y nutrigenética: la base genética de la nutriciónMario MoralesNoch keine Bewertungen
- Tema 1 ING GDokument3 SeitenTema 1 ING GMARIA DEL CARMEN MOLINA HIGUERASNoch keine Bewertungen
- TP Integrador 9Dokument1 SeiteTP Integrador 9Kevin Andres Z.Noch keine Bewertungen
- TALLER 6 - Transcripción IDokument6 SeitenTALLER 6 - Transcripción ILina HernandezNoch keine Bewertungen
- Resumen de Regulacion A Nivel DNADokument11 SeitenResumen de Regulacion A Nivel DNALuis Antonio Perez AvitiaNoch keine Bewertungen
- p53 Presentacion ScribdDokument26 Seitenp53 Presentacion ScribdFelipe RíosNoch keine Bewertungen
- MicrosateliteDokument7 SeitenMicrosateliteRicardo SotalìnNoch keine Bewertungen
- Organización y Regulación de La Transcripción GenéticaDokument7 SeitenOrganización y Regulación de La Transcripción GenéticaorutraodnamraNoch keine Bewertungen
- Regulacion Expresion GenicaDokument13 SeitenRegulacion Expresion Genicamanuel alejandro copairaNoch keine Bewertungen
- CTP 2 Segundo PDokument6 SeitenCTP 2 Segundo PMARIA PAULA MORALES RAMIREZNoch keine Bewertungen
- Tema 7 - Regulaci N de La Expresi N G NicaDokument8 SeitenTema 7 - Regulaci N de La Expresi N G NicaJON LEGUINANoch keine Bewertungen
- Guía de Estudio Regulación GénicaDokument23 SeitenGuía de Estudio Regulación GénicaNicolas Patricio Sitja100% (1)
- Transcripción Del ADNDokument17 SeitenTranscripción Del ADNdarwin plazaNoch keine Bewertungen
- 7 Transcripción (Modo de Compatibilidad)Dokument63 Seiten7 Transcripción (Modo de Compatibilidad)Villalòn Saldaña ChokyNoch keine Bewertungen
- 3 Transcripcion EucariontesDokument15 Seiten3 Transcripcion EucariontesMelissa AlonsoNoch keine Bewertungen
- La Celula (Organelas y Transcripción)Dokument50 SeitenLa Celula (Organelas y Transcripción)Maria LauraNoch keine Bewertungen
- El ADN, la molécula de la vida y su código genéticoDokument7 SeitenEl ADN, la molécula de la vida y su código genéticoangel azNoch keine Bewertungen
- TranscripcionDokument4 SeitenTranscripcionmafermoralesv1Noch keine Bewertungen
- Transcripcion MonzacolDokument6 SeitenTranscripcion MonzacolCarlaNoch keine Bewertungen
- Análisis Del DNA-IDokument11 SeitenAnálisis Del DNA-IMiguel Ángel Martínez MartínNoch keine Bewertungen
- El Código GenéticoDokument4 SeitenEl Código GenéticoWilmer NuñezNoch keine Bewertungen
- Transcripción en Eucariotas: Etapas, Promotores y Factores de Transcripción.Dokument4 SeitenTranscripción en Eucariotas: Etapas, Promotores y Factores de Transcripción.Arturo BenavidesNoch keine Bewertungen
- Caso 4 BCMDokument5 SeitenCaso 4 BCMCarolina Rivera ReyNoch keine Bewertungen
- TALLER Expresion GenicaDokument4 SeitenTALLER Expresion GenicaValentina Marin PeñatesNoch keine Bewertungen
- Genes, estructura y función molecularDokument26 SeitenGenes, estructura y función molecularCecilia Quiroga De DavilaNoch keine Bewertungen
- TALLER DE PREGUNTAS Biologia Molecular 3Dokument5 SeitenTALLER DE PREGUNTAS Biologia Molecular 3Diego RodriguezNoch keine Bewertungen
- Epigenética Del Cáncer Llega A La Oncología ConvencionalDokument15 SeitenEpigenética Del Cáncer Llega A La Oncología ConvencionalCamilo AriasNoch keine Bewertungen
- Presentación Universitaria Biología Sanitaria Fotográfico AzulDokument28 SeitenPresentación Universitaria Biología Sanitaria Fotográfico Azulmarylin sanchezNoch keine Bewertungen
- Grupo Nº1 - SEMANA13Dokument4 SeitenGrupo Nº1 - SEMANA13Ale VeraNoch keine Bewertungen
- Semana 3 - Auditoria de La SaludDokument3 SeitenSemana 3 - Auditoria de La SaludAle VeraNoch keine Bewertungen
- Medidas de Prevención Covid-19Dokument40 SeitenMedidas de Prevención Covid-19Ale Vera100% (1)
- Resumen de Infiltracion Anestesica LocalDokument3 SeitenResumen de Infiltracion Anestesica LocalAle VeraNoch keine Bewertungen
- Foro CulturaDokument2 SeitenForo CulturaAle VeraNoch keine Bewertungen
- Importancia de la medicina del trabajo y relaciones entre salud y trabajoDokument3 SeitenImportancia de la medicina del trabajo y relaciones entre salud y trabajoAle VeraNoch keine Bewertungen
- Peligros y RiesgosDokument2 SeitenPeligros y RiesgosAle VeraNoch keine Bewertungen
- Grupo Nº1 - SEMANA12Dokument5 SeitenGrupo Nº1 - SEMANA12Ale VeraNoch keine Bewertungen
- Grupo Nº1 - SEMANA11Dokument3 SeitenGrupo Nº1 - SEMANA11Ale VeraNoch keine Bewertungen
- GraficosDokument5 SeitenGraficosAle VeraNoch keine Bewertungen
- Sistema Nervioso CentralDokument1 SeiteSistema Nervioso CentralAle VeraNoch keine Bewertungen
- Lista Farmacos A Realizar Ficha Terapéutica Ii UnidadDokument2 SeitenLista Farmacos A Realizar Ficha Terapéutica Ii UnidadAle VeraNoch keine Bewertungen
- Gobierno Del Perú PDFDokument1 SeiteGobierno Del Perú PDFAle VeraNoch keine Bewertungen
- Bio Esta Di SticaDokument2 SeitenBio Esta Di SticaAle VeraNoch keine Bewertungen
- Alimentos para Aumentar El Colesterol HDLDokument4 SeitenAlimentos para Aumentar El Colesterol HDLAle Vera100% (1)
- Sindrome de GuillainDokument2 SeitenSindrome de GuillainAle VeraNoch keine Bewertungen
- BIOFISICA CLASE 13 2018 II TERMODINAMICA Termodin 1Dokument14 SeitenBIOFISICA CLASE 13 2018 II TERMODINAMICA Termodin 1Ale VeraNoch keine Bewertungen
- MALNUTRICIÓNDokument22 SeitenMALNUTRICIÓNAle VeraNoch keine Bewertungen
- Temperaturas ExtremasDokument17 SeitenTemperaturas ExtremasAle VeraNoch keine Bewertungen
- Experimento de MareyDokument12 SeitenExperimento de MareyDanielAndreeSalazarAlvarado0% (1)
- Seminario 7Dokument17 SeitenSeminario 7Ale VeraNoch keine Bewertungen
- Metodologia de La InvestigacionDokument3 SeitenMetodologia de La InvestigacionAle VeraNoch keine Bewertungen
- Informe de BioquimicaDokument2 SeitenInforme de BioquimicaAle VeraNoch keine Bewertungen
- Clasificación de Los Diferentes Tipos de Obesidad ExistentesDokument5 SeitenClasificación de Los Diferentes Tipos de Obesidad ExistentesAle VeraNoch keine Bewertungen
- Trata Mien ToDokument2 SeitenTrata Mien ToAle VeraNoch keine Bewertungen
- Determinacion de DesnutricionDokument14 SeitenDeterminacion de DesnutricionAle VeraNoch keine Bewertungen
- Actividad ExperimentalDokument11 SeitenActividad ExperimentalAle Vera100% (1)
- Informe Del 1 - 7Dokument13 SeitenInforme Del 1 - 7Ale VeraNoch keine Bewertungen
- MarasmoDokument5 SeitenMarasmoAle VeraNoch keine Bewertungen
- Sindrome MetabólicoDokument7 SeitenSindrome MetabólicoAle VeraNoch keine Bewertungen
- 5 Semana BiologiaDokument5 Seiten5 Semana BiologiaErnestoLazaroGomeroNoch keine Bewertungen
- Práctica #03Dokument3 SeitenPráctica #03Carlos Llicán CenturiónNoch keine Bewertungen
- Aminoácidos y proteínas: Fundamentos biológicos de la medicinaDokument3 SeitenAminoácidos y proteínas: Fundamentos biológicos de la medicinacatailnaNoch keine Bewertungen
- Importancia Biológica Del MetabolismoDokument2 SeitenImportancia Biológica Del MetabolismomonipolyNoch keine Bewertungen
- Glosario Biologia MolecularDokument10 SeitenGlosario Biologia MolecularGrupo A InnovaciónNoch keine Bewertungen
- Impronta GenómicaDokument2 SeitenImpronta GenómicaDavid Herrera100% (1)
- Factores de La Coagulacion PDFDokument2 SeitenFactores de La Coagulacion PDFJoshNoch keine Bewertungen
- Cuestionario de FarmacologiaDokument3 SeitenCuestionario de FarmacologiaJaime San100% (2)
- Fisiología Del Metabolismo de La ProteínasDokument19 SeitenFisiología Del Metabolismo de La ProteínasReinaldo Rodriguez Guerrero100% (5)
- MicroarreglosDokument21 SeitenMicroarreglosAlejandraNoch keine Bewertungen
- Retículo EndoplasmáticoDokument3 SeitenRetículo EndoplasmáticoLulú Frank OrtizNoch keine Bewertungen
- Examen de Periodo 1Dokument3 SeitenExamen de Periodo 1Alexander chaucanesNoch keine Bewertungen
- CAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoDokument91 SeitenCAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoAlexis AlonsoNoch keine Bewertungen
- Usos de Reactivos en La Extracción de ADN y ElectroforesisDokument2 SeitenUsos de Reactivos en La Extracción de ADN y Electroforesiskaritho sanchezNoch keine Bewertungen
- ESTUDIO DE LA PANDEMIA y ANÁLISIS CIENTÍFICO INDEPENDIENTE Dr. Sergio J. Pérez OliveroDokument2.290 SeitenESTUDIO DE LA PANDEMIA y ANÁLISIS CIENTÍFICO INDEPENDIENTE Dr. Sergio J. Pérez OliveroYankaNoch keine Bewertungen
- T18 Ingenieria Genética EdomingoDokument30 SeitenT18 Ingenieria Genética Edomingomanuroldaan 12Noch keine Bewertungen
- Factores de Crecimiento y Señalización en CáncerDokument7 SeitenFactores de Crecimiento y Señalización en CáncerRODRIGO YAEL P CABANILLAS100% (1)
- Inhibición enzimática competitiva y no competitivaDokument2 SeitenInhibición enzimática competitiva y no competitivaDavid Mauricio CentenoNoch keine Bewertungen
- Síndrome Letal Overo BlancoDokument32 SeitenSíndrome Letal Overo BlancocasanchezisNoch keine Bewertungen
- BBM1 U1 Atr MagpDokument4 SeitenBBM1 U1 Atr MagpAngeles GrajalesNoch keine Bewertungen
- Mecanismos de Acción de Las QuinonasDokument2 SeitenMecanismos de Acción de Las QuinonasDamián AlexanderNoch keine Bewertungen
- Biolementos y BiomoleculasDokument13 SeitenBiolementos y BiomoleculasCrisangel Sequera50% (2)
- División Celular y Reproduccion CelularDokument15 SeitenDivisión Celular y Reproduccion CelularRocio Pilar Parra SkinnerNoch keine Bewertungen
- Trabajo Sobre APOPTOSIS Con Retroalimentación Del Docente.Dokument4 SeitenTrabajo Sobre APOPTOSIS Con Retroalimentación Del Docente.Leonardo Rachen RodriguezNoch keine Bewertungen
- Biologia 25 Tarea 3 AdrianaVelasquezDokument8 SeitenBiologia 25 Tarea 3 AdrianaVelasquezAdriana Del PilarNoch keine Bewertungen
- ¿La Estructura ImportaDokument1 Seite¿La Estructura ImportaCris CidLNoch keine Bewertungen
- TransSinápticaNeurotransmisoresDokument3 SeitenTransSinápticaNeurotransmisoresChristian Correa VerdeNoch keine Bewertungen
- Objetivos BiocaDokument286 SeitenObjetivos BiocaDaniel Kamarov FedchenkoNoch keine Bewertungen
- El ciclo celular: G1, S, G2 y MDokument4 SeitenEl ciclo celular: G1, S, G2 y MCami QeNoch keine Bewertungen
- Inhibidores enzimáticos U. San AngelDokument45 SeitenInhibidores enzimáticos U. San AngelMarysol MartinezNoch keine Bewertungen