Das könnte Ihnen auch gefallen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
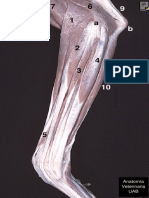
- Anatomía Veterinaria UABDokument1 SeiteAnatomía Veterinaria UABomarcvetNoch keine Bewertungen
- Anatomía Veterinaria UABDokument1 SeiteAnatomía Veterinaria UABomarcvetNoch keine Bewertungen
- Anatomía Veterinaria UABDokument1 SeiteAnatomía Veterinaria UABomarcvetNoch keine Bewertungen
- Anatomía Veterinaria UABDokument1 SeiteAnatomía Veterinaria UABomarcvetNoch keine Bewertungen
- Glucogeno PDFDokument24 SeitenGlucogeno PDFomarcvetNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1091)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- 50 Cross-Connection Questions, Answers & IllustrationsDokument12 Seiten50 Cross-Connection Questions, Answers & IllustrationsCherokeeMetro100% (1)
- Review On Factors Which Affect Coffee (Coffea Arabica L.) Quality in South Western, EthiopiaDokument8 SeitenReview On Factors Which Affect Coffee (Coffea Arabica L.) Quality in South Western, EthiopiaErin WalkerNoch keine Bewertungen
- Sustainable DevelopmentDokument33 SeitenSustainable DevelopmentMa Rosario H. Mallari100% (1)
- Chemistry Perfumes Your Daily LifeDokument6 SeitenChemistry Perfumes Your Daily Lifeฝน เมนโดซาNoch keine Bewertungen
- Brunei Imports 3,003 Tons of Jasmine Rice From Cambodia: TOP Contents - Tailored For YOUDokument16 SeitenBrunei Imports 3,003 Tons of Jasmine Rice From Cambodia: TOP Contents - Tailored For YOUMujahid AliNoch keine Bewertungen
- Chen Et Al. (2017) - Effects of Terracing Practices On Water Erosion Control in ChinaDokument35 SeitenChen Et Al. (2017) - Effects of Terracing Practices On Water Erosion Control in ChinaDemetrio Angelo Lama IsminioNoch keine Bewertungen
- Final Copy 558 This Is A Mini ProjectDokument29 SeitenFinal Copy 558 This Is A Mini ProjectGurusreenuNoch keine Bewertungen
- Cooperative Finance in Developing EconomiesDokument282 SeitenCooperative Finance in Developing EconomiesSourav DharNoch keine Bewertungen
- Determination of Aflatoxins H P L C ColumnsDokument6 SeitenDetermination of Aflatoxins H P L C ColumnsIvanaColinNoch keine Bewertungen
- 34 Socio Economic Review EnglishDokument234 Seiten34 Socio Economic Review EnglishjarikhatriNoch keine Bewertungen
- Mothbean Production: Dr. M M Sharma Moth Breeder SK Rau, BikanerDokument9 SeitenMothbean Production: Dr. M M Sharma Moth Breeder SK Rau, Bikanersatya6a9246Noch keine Bewertungen
- Cactus: D. in The Arid and Hot RegionDokument4 SeitenCactus: D. in The Arid and Hot RegionDamaraji Nurwidhi Anggara100% (1)
- 2.for Rural LOw Volume Traffic Rigid Pavement DesignDokument47 Seiten2.for Rural LOw Volume Traffic Rigid Pavement DesignTarak A Positive100% (2)
- Itihass 11 VIDokument138 SeitenItihass 11 VIsreyrnNoch keine Bewertungen
- Distribution and Habitat: Jasminum Sambac Jasminum Mesnyi Jasminum AngustifoliumDokument1 SeiteDistribution and Habitat: Jasminum Sambac Jasminum Mesnyi Jasminum AngustifoliumpriyankaNoch keine Bewertungen
- 10 Tips For Mowing Your Lawn PerfectlyDokument2 Seiten10 Tips For Mowing Your Lawn PerfectlyLifestyle LawnsNoch keine Bewertungen
- Presentation 3 History of PTCDokument34 SeitenPresentation 3 History of PTCSunita RanawatNoch keine Bewertungen
- 2010 Pulse Processing Functionality and Application Litera.Dokument145 Seiten2010 Pulse Processing Functionality and Application Litera.abid222Noch keine Bewertungen
- Broom GrassDokument16 SeitenBroom GrassEddie FetalveroNoch keine Bewertungen
- 13th and 14th Century SocietiesDokument6 Seiten13th and 14th Century SocietiesAnuraj TirthankarNoch keine Bewertungen
- Shattered PlaytestDokument98 SeitenShattered PlaytestKen100% (2)
- PROPERTY - TITLE V and Beyond List of Cases Possession: Director of Lands Vs IAC, May 1992Dokument7 SeitenPROPERTY - TITLE V and Beyond List of Cases Possession: Director of Lands Vs IAC, May 1992ZahraMinaNoch keine Bewertungen
- Agriculture and Chemical CompaniesDokument8 SeitenAgriculture and Chemical CompaniesRajiv RanjanNoch keine Bewertungen
- What Is Human Settlement?: Types of Human SettlementsDokument25 SeitenWhat Is Human Settlement?: Types of Human Settlementszandra mallariNoch keine Bewertungen
- November 4, 2016 Strathmore TimesDokument28 SeitenNovember 4, 2016 Strathmore TimesStrathmore TimesNoch keine Bewertungen
- Journal of Economic Education: Deo Leko Pudaka, Rusdarti, P. Eko PrasetyoDokument8 SeitenJournal of Economic Education: Deo Leko Pudaka, Rusdarti, P. Eko PrasetyoDedy Asep IrawanNoch keine Bewertungen
- Sumario Executivo Iniciativa Clima e Desenvolvimento ENGLISHDokument16 SeitenSumario Executivo Iniciativa Clima e Desenvolvimento ENGLISHMichele CottaNoch keine Bewertungen
- Adaptation in PlantsDokument3 SeitenAdaptation in PlantsAbhisek SarkarNoch keine Bewertungen
- Exercise Unit 2Dokument8 SeitenExercise Unit 2Hân HoàngNoch keine Bewertungen
- Experience of An FSOE ExamineeDokument2 SeitenExperience of An FSOE ExamineeReddick RoseNoch keine Bewertungen