Das könnte Ihnen auch gefallen
- Nutraceuticals and Natural Product Derivatives: Disease Prevention & Drug DiscoveryVon EverandNutraceuticals and Natural Product Derivatives: Disease Prevention & Drug DiscoveryNoch keine Bewertungen
- Antifatigue and Antioxidant Activity of Alcoh - 2011 - Journal of Traditional AnDokument5 SeitenAntifatigue and Antioxidant Activity of Alcoh - 2011 - Journal of Traditional Ankuya batokNoch keine Bewertungen
- 2013 Article 1220Dokument11 Seiten2013 Article 1220MeLissa AymNoch keine Bewertungen
- 1 s2.0 S2405654520300032 MainDokument9 Seiten1 s2.0 S2405654520300032 MainRosalin nathNoch keine Bewertungen
- 18 PDFDokument6 Seiten18 PDFТатьяна СаенкоNoch keine Bewertungen
- 2021 Alleviation of Diabetes-Induced Hepatotoxicity by Date Palm Hydroalcoholic Extract in Rat Model - A Biochemical, Immunohistochemical and Stereological StudyDokument10 Seiten2021 Alleviation of Diabetes-Induced Hepatotoxicity by Date Palm Hydroalcoholic Extract in Rat Model - A Biochemical, Immunohistochemical and Stereological StudyemanNoch keine Bewertungen
- Propolis Hepatoprotector Effect On Liver Damage of White Mice Induced by Valproic AcidDokument12 SeitenPropolis Hepatoprotector Effect On Liver Damage of White Mice Induced by Valproic AcidHarrizul RivaiNoch keine Bewertungen
- Intense and Exhaustive Exercise Induce Oxidative Stress in Skeletal MuscleDokument4 SeitenIntense and Exhaustive Exercise Induce Oxidative Stress in Skeletal MuscleRoy WilsonNoch keine Bewertungen
- Black Seed 5Dokument8 SeitenBlack Seed 5abdulNoch keine Bewertungen
- Hepatoprotective and Antioxidants Activity of Ethanolic Extract of Cuscutta Reflexa RoxbDokument6 SeitenHepatoprotective and Antioxidants Activity of Ethanolic Extract of Cuscutta Reflexa RoxbIOSR Journal of PharmacyNoch keine Bewertungen
- 1 s2.0 S175646461930622X MainDokument10 Seiten1 s2.0 S175646461930622X MainEli SahirohNoch keine Bewertungen
- Effects of Lipoic Acid Supplementation On Rat Brain TissueDokument8 SeitenEffects of Lipoic Acid Supplementation On Rat Brain Tissueqiang liNoch keine Bewertungen
- Research ArticleDokument9 SeitenResearch ArticleElfiaNeswitaNoch keine Bewertungen
- Effect of Purple Sweet Potato Ipomoea Ba 24953e1eDokument8 SeitenEffect of Purple Sweet Potato Ipomoea Ba 24953e1e1005 Siti Ainunnisa Lingga WinariNoch keine Bewertungen
- Research Article Moringa Oleifera in Rats Subjected ToDokument10 SeitenResearch Article Moringa Oleifera in Rats Subjected ToYaniNoch keine Bewertungen
- TLC Bioautography-Guided Isolation of AntioxidantsDokument6 SeitenTLC Bioautography-Guided Isolation of AntioxidantsArieNoch keine Bewertungen
- Purification and Identification of Antioxidant Peptides From Egg White Protein HydrolysateDokument10 SeitenPurification and Identification of Antioxidant Peptides From Egg White Protein Hydrolysateمصطفى محمد عبدالمنعمNoch keine Bewertungen
- Ahn2014 PDFDokument6 SeitenAhn2014 PDFVenny AgustinNoch keine Bewertungen
- Ngu Gia Bi Gai-Acanthopanax SenticosusDokument6 SeitenNgu Gia Bi Gai-Acanthopanax SenticosusMai Anh NguyễnNoch keine Bewertungen
- Food Chemistry: Hui-Yin Chen, Gow-Chin YenDokument9 SeitenFood Chemistry: Hui-Yin Chen, Gow-Chin YenGREESTYNoch keine Bewertungen
- Research Article Schisandra Chinensis, Artemisia Capillaris, and Aloe BarbadensisDokument11 SeitenResearch Article Schisandra Chinensis, Artemisia Capillaris, and Aloe Barbadensismuhamad patkurohmanNoch keine Bewertungen
- Effect of The Long-Term Intake of An Egg PDFDokument7 SeitenEffect of The Long-Term Intake of An Egg PDFMohammed HussienNoch keine Bewertungen
- Hepatoprotective Activity of Homalomena Aromatica Against CCL4 Induced Liver Injury in Swiss Albino MiceDokument8 SeitenHepatoprotective Activity of Homalomena Aromatica Against CCL4 Induced Liver Injury in Swiss Albino MiceIJAR JOURNALNoch keine Bewertungen
- Food and Chemical Toxicology: Hussain Zeashan, G. Amresh, Satyawan Singh, Chandana Venkateswara RaoDokument5 SeitenFood and Chemical Toxicology: Hussain Zeashan, G. Amresh, Satyawan Singh, Chandana Venkateswara RaodanilriosNoch keine Bewertungen
- The Effects of Angelica Keiskei Koidz On The Expression of Antioxidant Enzymes Related To Lipid Profiles in Rats Fed A High Fat DietDokument7 SeitenThe Effects of Angelica Keiskei Koidz On The Expression of Antioxidant Enzymes Related To Lipid Profiles in Rats Fed A High Fat DietAriWahyuNoch keine Bewertungen
- Pharmacology, Biochemistry and BehaviorDokument6 SeitenPharmacology, Biochemistry and BehaviorFerviferrazNoch keine Bewertungen
- Possible Mechanism of Plumbago Zeylanica in Prevention of Hepatic Dammage in Wistar RatDokument8 SeitenPossible Mechanism of Plumbago Zeylanica in Prevention of Hepatic Dammage in Wistar RatAmit patelNoch keine Bewertungen
- 9 PDFDokument7 Seiten9 PDFfaikaNoch keine Bewertungen
- Food and Chemical Toxicology: Bayram Yurt, Ismail CelikDokument6 SeitenFood and Chemical Toxicology: Bayram Yurt, Ismail CelikGÖKHAN DURMAZNoch keine Bewertungen
- BT 26 225Dokument17 SeitenBT 26 225Seção de Beneficiamento Fazenda da AeronáuticaNoch keine Bewertungen
- Present As IDokument9 SeitenPresent As Iaini rahmawatiNoch keine Bewertungen
- Ananingsih 2013Dokument11 SeitenAnaningsih 2013Phung Vo Hung PhatNoch keine Bewertungen
- Antioxidative Potential of Cynodon DactyloDokument5 SeitenAntioxidative Potential of Cynodon Dactylochaitanya gNoch keine Bewertungen
- 154 PDFDokument5 Seiten154 PDFRusniaty AbdulrahmanNoch keine Bewertungen
- Liu - 2011 - The Antioxidant and Free-Radical Scavenging Activities of Extract and Fractions PDFDokument9 SeitenLiu - 2011 - The Antioxidant and Free-Radical Scavenging Activities of Extract and Fractions PDFAndika Yuli HeryantoNoch keine Bewertungen
- Food Research International: Laura E. Gayosso-García Sancho, Elhadi M. Yahia, Gustavo Adolfo González-AguilarDokument8 SeitenFood Research International: Laura E. Gayosso-García Sancho, Elhadi M. Yahia, Gustavo Adolfo González-AguilarBEN DUNCAN MALAGA ESPICHANNoch keine Bewertungen
- Separation and Identification of Antioxidant Peptides From Foxtail Millet (Setaria Italica) Prolamins Enzymatic HydrolysateDokument35 SeitenSeparation and Identification of Antioxidant Peptides From Foxtail Millet (Setaria Italica) Prolamins Enzymatic HydrolysateDEINY MARYELI CORDOBA CERONNoch keine Bewertungen
- 1 s2.0 S0032579119310156 MainDokument7 Seiten1 s2.0 S0032579119310156 MainFAYYADH ZAIDAAN AKHIRU ICHYA PUTRANoch keine Bewertungen
- Structure characterization and antitumor activity of an α β-glucan polysaccharideDokument7 SeitenStructure characterization and antitumor activity of an α β-glucan polysaccharideMarcela LaraNoch keine Bewertungen
- Food and Chemical Toxicology: Engin Celep, Ahmet Aydın, Hasan Kırmızıbekmez, Erdem YesiladaDokument8 SeitenFood and Chemical Toxicology: Engin Celep, Ahmet Aydın, Hasan Kırmızıbekmez, Erdem YesiladaDivaruzNoch keine Bewertungen
- Chemical Composition of Five Wild Edible Mushrooms CollectedDokument7 SeitenChemical Composition of Five Wild Edible Mushrooms Collectedela.sofiaNoch keine Bewertungen
- P. 257-265Dokument9 SeitenP. 257-265Aisyaah imranNoch keine Bewertungen
- 1 s2.0 S075333220700234X MainDokument8 Seiten1 s2.0 S075333220700234X MainManar HelmyNoch keine Bewertungen
- 1 s2.0 S1756464617302876 MainDokument10 Seiten1 s2.0 S1756464617302876 MainHussain AhmadNoch keine Bewertungen
- Neuropharmacological Profile of Eclipta Alba (Linn.) HasskDokument9 SeitenNeuropharmacological Profile of Eclipta Alba (Linn.) Hassktanvir.radiant officerNoch keine Bewertungen
- Gastroprotective Effect of The Traditional Herbal Medicine, Sipjeondaebo-Tang Water Extract, Against Ethanol-Induced Gastric Mucosal InjuryDokument7 SeitenGastroprotective Effect of The Traditional Herbal Medicine, Sipjeondaebo-Tang Water Extract, Against Ethanol-Induced Gastric Mucosal InjuryPhilip ScofieldNoch keine Bewertungen
- Putative Benefits of Microalgal Astaxanthin On Exercise and Human HealthDokument7 SeitenPutative Benefits of Microalgal Astaxanthin On Exercise and Human HealthRaziel Alvarez RebolloNoch keine Bewertungen
- Protective Effect of Allicin Against Acrylamide-Induced Hepatocyte Damage in Vitro and in Vivo1Dokument7 SeitenProtective Effect of Allicin Against Acrylamide-Induced Hepatocyte Damage in Vitro and in Vivo1PostNL ServiceNoch keine Bewertungen
- Food and Chemical Toxicology: Lijun Sun, Jianbao Zhang, Xiaoyun Lu, Liyu Zhang, Yali ZhangDokument8 SeitenFood and Chemical Toxicology: Lijun Sun, Jianbao Zhang, Xiaoyun Lu, Liyu Zhang, Yali ZhangSri LakshmiNoch keine Bewertungen
- Spiso 11 PDFDokument15 SeitenSpiso 11 PDFGuaguanconNoch keine Bewertungen
- 12 - AsurDokument8 Seiten12 - AsurFujiko SaavedraNoch keine Bewertungen
- Preliminary Clinical Evaluation of Toxicity and Efficacy of A New Astaxanthin-Rich Haematococcus Pluvialis ExtractDokument5 SeitenPreliminary Clinical Evaluation of Toxicity and Efficacy of A New Astaxanthin-Rich Haematococcus Pluvialis ExtractEva Pa'e ONoch keine Bewertungen
- Abst FlavonoidesDokument7 SeitenAbst Flavonoidesdeysy galvan ayalaNoch keine Bewertungen
- Efficacy and Safety of Chinese Black Tea (Pu-Ehr) Extract in Healthy and Hypercholesterolemic SubjectsDokument11 SeitenEfficacy and Safety of Chinese Black Tea (Pu-Ehr) Extract in Healthy and Hypercholesterolemic SubjectsdanielwongtsNoch keine Bewertungen
- 005 Sudirman2021 - Hippocampus Kuda Protein Hydrolysate Improves Male Reproductive - RedDokument9 Seiten005 Sudirman2021 - Hippocampus Kuda Protein Hydrolysate Improves Male Reproductive - RedSabri SudirmanNoch keine Bewertungen
- Gastroprotective Effects of Ozonized Sunflower Oil-5dec06Dokument6 SeitenGastroprotective Effects of Ozonized Sunflower Oil-5dec06zyz777Noch keine Bewertungen
- Food and Chemical Toxicology: Article InfoDokument7 SeitenFood and Chemical Toxicology: Article InfoNathalia MojicaNoch keine Bewertungen
- A novel oligosaccharide isolated from Hericium erinaceus and its protection against LPS-induced Caco-2 cells via the TLR4 - NF-κB pathwayDokument9 SeitenA novel oligosaccharide isolated from Hericium erinaceus and its protection against LPS-induced Caco-2 cells via the TLR4 - NF-κB pathwaymalvina.sekolonik911Noch keine Bewertungen
- Are Polyphenols Antioxidants or Pro-Oxidants What Do We LearnDokument6 SeitenAre Polyphenols Antioxidants or Pro-Oxidants What Do We LearnDiana Maria Amaya CruzNoch keine Bewertungen
- Histolological Effects of Pomesteen Power Supplement On The Liver of Adult Wistar RatsDokument9 SeitenHistolological Effects of Pomesteen Power Supplement On The Liver of Adult Wistar RatsSteven JonesNoch keine Bewertungen
- Daftar PustakaDokument3 SeitenDaftar PustakaTiara Grhanesia DenashuryaNoch keine Bewertungen
- Morning Report 25 NovemberDokument57 SeitenMorning Report 25 NovemberTiara Grhanesia DenashuryaNoch keine Bewertungen
- Morning Report 21 November 2014Dokument54 SeitenMorning Report 21 November 2014Tiara Grhanesia DenashuryaNoch keine Bewertungen
- Morning Report 24 November 2014Dokument53 SeitenMorning Report 24 November 2014Tiara Grhanesia DenashuryaNoch keine Bewertungen
- Exercise Is A Type of Physical Activity Consisting of Planned, Structured, and Repetitive BodilyDokument3 SeitenExercise Is A Type of Physical Activity Consisting of Planned, Structured, and Repetitive BodilyTiara Grhanesia DenashuryaNoch keine Bewertungen
- Template Morning Report FK UGMDokument55 SeitenTemplate Morning Report FK UGMTiara Grhanesia DenashuryaNoch keine Bewertungen
- ADokument2 SeitenATiara Grhanesia DenashuryaNoch keine Bewertungen
- Referat Sarkopenia Pada GeriatriDokument2 SeitenReferat Sarkopenia Pada GeriatriTiara Grhanesia DenashuryaNoch keine Bewertungen
- Answer Key For Reading PracticeDokument1 SeiteAnswer Key For Reading PracticeTiara Grhanesia DenashuryaNoch keine Bewertungen
- Cara Baca CT ScanDokument180 SeitenCara Baca CT ScanTiara Grhanesia DenashuryaNoch keine Bewertungen
- Cara Baca CT ScanDokument180 SeitenCara Baca CT ScanTiara Grhanesia DenashuryaNoch keine Bewertungen
- Peptic Ulcer Bleeding Risk. The Role of Helicobacter: Pylori Infection in NSAID/Low-Dose Aspirin UsersDokument1 SeitePeptic Ulcer Bleeding Risk. The Role of Helicobacter: Pylori Infection in NSAID/Low-Dose Aspirin UsersTiara Grhanesia DenashuryaNoch keine Bewertungen
- Acls PDFDokument53 SeitenAcls PDFKeez HaruNoch keine Bewertungen
- Harris SDokument2 SeitenHarris STiara Grhanesia DenashuryaNoch keine Bewertungen
- Fave Rio 2014Dokument8 SeitenFave Rio 2014Tiara Grhanesia DenashuryaNoch keine Bewertungen
- Hyponatremia in Children With Acute Respiratory Infections: A ReappraisalDokument6 SeitenHyponatremia in Children With Acute Respiratory Infections: A ReappraisalTiara Grhanesia DenashuryaNoch keine Bewertungen
- Accepted Manuscript: YprrvDokument27 SeitenAccepted Manuscript: YprrvTiara Grhanesia DenashuryaNoch keine Bewertungen
- Protocols For Hyperlactatemia Induction in The Lactate Minimum Test Adapted To Swimming RatsDokument5 SeitenProtocols For Hyperlactatemia Induction in The Lactate Minimum Test Adapted To Swimming RatsTiara Grhanesia DenashuryaNoch keine Bewertungen
- JGJDokument55 SeitenJGJTiara Grhanesia DenashuryaNoch keine Bewertungen
- The God Helmet PDFDokument5 SeitenThe God Helmet PDFTiara Grhanesia DenashuryaNoch keine Bewertungen
- Chat Kin 2016Dokument8 SeitenChat Kin 2016Tiara Grhanesia DenashuryaNoch keine Bewertungen
- Guideline For Hypertensive Emergency Treatment PDFDokument4 SeitenGuideline For Hypertensive Emergency Treatment PDFTiara Grhanesia DenashuryaNoch keine Bewertungen
- 18 Arias 018Dokument24 Seiten18 Arias 018Tiara Grhanesia DenashuryaNoch keine Bewertungen
- Oral Ondansetron For Gastroenteritis in A Pediatric Emergency DepartmentDokument8 SeitenOral Ondansetron For Gastroenteritis in A Pediatric Emergency DepartmentTiara Grhanesia DenashuryaNoch keine Bewertungen
- I. Patient IdentityDokument20 SeitenI. Patient IdentityTiara Grhanesia DenashuryaNoch keine Bewertungen
- Cah 01 10 Ts DiarrDokument14 SeitenCah 01 10 Ts DiarrTiara Grhanesia DenashuryaNoch keine Bewertungen
- Acute Diare WHO 2005Dokument50 SeitenAcute Diare WHO 2005Boby Abdul RahmanNoch keine Bewertungen
- 2016 Acute Diarrhea PDFDokument21 Seiten2016 Acute Diarrhea PDFTiara Grhanesia DenashuryaNoch keine Bewertungen
- Bohu 2014Dokument7 SeitenBohu 2014Tiara Grhanesia DenashuryaNoch keine Bewertungen
- Anti Icing T4Dokument13 SeitenAnti Icing T4Mansooooor12345Noch keine Bewertungen
- Fish Smoking ProceduresDokument42 SeitenFish Smoking Proceduresanegbe1199100% (1)
- Cooling System Report (Automotive Tech.)Dokument5 SeitenCooling System Report (Automotive Tech.)Mohamed AhmedNoch keine Bewertungen
- The Light BulbDokument4 SeitenThe Light BulbJonas SarmientoNoch keine Bewertungen
- Bahrain O&g Facts and Figurs 2012 enDokument60 SeitenBahrain O&g Facts and Figurs 2012 ennjava1978Noch keine Bewertungen
- DANAIS MT II CL 300 ENDokument30 SeitenDANAIS MT II CL 300 ENLasombra DelPatoNoch keine Bewertungen
- DEC Proposal For OU7Dokument9 SeitenDEC Proposal For OU7MonicaNoch keine Bewertungen
- Unit I (IC Engines)Dokument13 SeitenUnit I (IC Engines)prasanthiNoch keine Bewertungen
- Gönen, Turan-Modern Power System Analysis-CRC Press (2013)Dokument47 SeitenGönen, Turan-Modern Power System Analysis-CRC Press (2013)Anand KalNoch keine Bewertungen
- SD7000 InstructionsDokument8 SeitenSD7000 InstructionsMarcelo CheloNoch keine Bewertungen
- Reliance - 10080-1-SS-PP-014 - REV2Dokument58 SeitenReliance - 10080-1-SS-PP-014 - REV2Blain86% (7)
- RBSA Indian Steel Industry AnalysisDokument45 SeitenRBSA Indian Steel Industry AnalysisVasiliy GuryevNoch keine Bewertungen
- Donnelly 2019 - Nat Methods - Protein Analysis MSDokument8 SeitenDonnelly 2019 - Nat Methods - Protein Analysis MSchuvanessNoch keine Bewertungen
- Concrete and Concrete Aggregates: Standard Terminology Relating ToDokument8 SeitenConcrete and Concrete Aggregates: Standard Terminology Relating ToV. ManilalNoch keine Bewertungen
- Tensar Basetex Technical Info (April 2003)Dokument4 SeitenTensar Basetex Technical Info (April 2003)sandycastleNoch keine Bewertungen
- Causes of KicksDokument14 SeitenCauses of KicksGhavban DavidNoch keine Bewertungen
- Corn Wet Milling Brochure 12209Dokument8 SeitenCorn Wet Milling Brochure 12209amanawatNoch keine Bewertungen
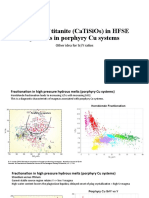
- The Role of Titanite (CaTiSiO ) in PCD ExplorationDokument9 SeitenThe Role of Titanite (CaTiSiO ) in PCD ExplorationMarco Augusto Robles AncajimaNoch keine Bewertungen
- Experimental Techniques For Measuring Sewing Needle TemperatureDokument8 SeitenExperimental Techniques For Measuring Sewing Needle TemperatureATLASNoch keine Bewertungen
- Extracting Bromine From Sea WaterDokument4 SeitenExtracting Bromine From Sea WaterRaja Ji100% (2)
- FoamMelt 200 Data SheetDokument2 SeitenFoamMelt 200 Data SheetNordson Adhesive Dispensing SystemsNoch keine Bewertungen
- BS 5950-1 1990Dokument126 SeitenBS 5950-1 1990Anoy100% (4)
- D2360 - 11 Standard Test Method For Trace Impurities in Monocyclic Aromatic Hydrocarbons by Gas Chromatography PDFDokument6 SeitenD2360 - 11 Standard Test Method For Trace Impurities in Monocyclic Aromatic Hydrocarbons by Gas Chromatography PDFJacques BlueqNoch keine Bewertungen
- Where There's Oil, Where There's Water, There's XylanDokument6 SeitenWhere There's Oil, Where There's Water, There's XylanWK SinnNoch keine Bewertungen
- Strain Hardening Rheology. Rheology Is The Science of Deformation. This Science Poses ADokument8 SeitenStrain Hardening Rheology. Rheology Is The Science of Deformation. This Science Poses Akhudhayer1970Noch keine Bewertungen
- Extra Solved Questions Class Ix Term II ChemistryDokument3 SeitenExtra Solved Questions Class Ix Term II Chemistrychhabra navdeep100% (1)
- EXEL - Insulating Rail Joints For Web PDFDokument4 SeitenEXEL - Insulating Rail Joints For Web PDFhariridkNoch keine Bewertungen
- IGCC For RefineryDokument20 SeitenIGCC For RefinerydensandsNoch keine Bewertungen
- Infrared Spectroscopy Absorption TableDokument7 SeitenInfrared Spectroscopy Absorption TableAmalinda Kharisma AdhaniNoch keine Bewertungen
- Parameter Study of Melt Spun Polypropylene Fibers by Centrifugal SpinningDokument16 SeitenParameter Study of Melt Spun Polypropylene Fibers by Centrifugal SpinningAnonymous PHCzwD8eAONoch keine Bewertungen