Das könnte Ihnen auch gefallen
- WordFamilySorts 1Dokument38 SeitenWordFamilySorts 1Karen simpsonNoch keine Bewertungen
- A Study On Welding Defects of Pressure VesselDokument24 SeitenA Study On Welding Defects of Pressure Vesseladamahmad1992100% (1)
- Brochure A35g A40g t4f en 22 20050840 DDokument20 SeitenBrochure A35g A40g t4f en 22 20050840 DRengga PratamaNoch keine Bewertungen
- Process Engineering ChiyodaDokument65 SeitenProcess Engineering ChiyodaSean Choi100% (10)
- Chemical Process Plant SafetyDokument37 SeitenChemical Process Plant Safetyvenky437Noch keine Bewertungen
- Poster SIL PDFDokument1 SeitePoster SIL PDFvgogulakrishnanNoch keine Bewertungen
- Cellular Automaton (Pl. Cellular Automata, Abbrev. CA) : Discrete Coupled Map LatticeDokument17 SeitenCellular Automaton (Pl. Cellular Automata, Abbrev. CA) : Discrete Coupled Map LatticeSimón Salgado MuñozNoch keine Bewertungen
- Project Proposal CarolingDokument7 SeitenProject Proposal CarolingAmparo Daniel Einstein D.Noch keine Bewertungen
- Caution!: Portable Digital Color Doppler Ultrasound SystemDokument177 SeitenCaution!: Portable Digital Color Doppler Ultrasound SystemDaniel Galindo100% (1)
- Free Convection (Formulae & Problems)Dokument15 SeitenFree Convection (Formulae & Problems)ananth2012Noch keine Bewertungen
- Return Material Authorization: 1/2 RMA Number Customer No. Purchase Order No. Sales Order DateDokument2 SeitenReturn Material Authorization: 1/2 RMA Number Customer No. Purchase Order No. Sales Order DateMichelle PadillaNoch keine Bewertungen
- BRKCCT-1002 Cisco Live Contact CenterDokument44 SeitenBRKCCT-1002 Cisco Live Contact CenterArun GuptaNoch keine Bewertungen
- Introduction to Non-Linear Mechanics. (AM-11), Volume 11Von EverandIntroduction to Non-Linear Mechanics. (AM-11), Volume 11Noch keine Bewertungen
- DuvalTriangles1 7-29mar2016Dokument68 SeitenDuvalTriangles1 7-29mar2016Jorgo BelloNoch keine Bewertungen
- Unit I - Discrete State-Variable Technique Q.No Questions: Scalar Adder IntegratorDokument184 SeitenUnit I - Discrete State-Variable Technique Q.No Questions: Scalar Adder IntegratorAgatha ChristieNoch keine Bewertungen
- Gas Plant Improves C3 Recovery With Lean Six Sigma ApproachDokument9 SeitenGas Plant Improves C3 Recovery With Lean Six Sigma ApproachganeshdhageNoch keine Bewertungen
- Cellular Automata & Its ApplicationsDokument57 SeitenCellular Automata & Its Applicationsvarun_bhatia2968076Noch keine Bewertungen
- Modelling With Cellular Automata (Slides)Dokument28 SeitenModelling With Cellular Automata (Slides)Mauro Sebastián Innocente100% (1)
- Cellular Automata: A Discrete View of The World: Joel L. SchiffDokument16 SeitenCellular Automata: A Discrete View of The World: Joel L. Schiffnada abdelrahmanNoch keine Bewertungen
- Cellular Automata, Entropy and Box - Coun4ng DimensionDokument28 SeitenCellular Automata, Entropy and Box - Coun4ng DimensionjamesNoch keine Bewertungen
- Cellular AutomataDokument27 SeitenCellular AutomataRahul Sharma100% (1)
- History: Original Concept of CA Is Most Strongly Associated With John Von Neumann - VonDokument11 SeitenHistory: Original Concept of CA Is Most Strongly Associated With John Von Neumann - Vondggil2Noch keine Bewertungen
- Paola Flocchini and Vladimir Cezar - Radial View of Continuous Cellular AutomataDokument19 SeitenPaola Flocchini and Vladimir Cezar - Radial View of Continuous Cellular AutomataldkgfdNoch keine Bewertungen
- Alexey Chilikov, Alexey Zhukov, Alexey Verkhovsky. The Research of Reversible Cellular Automata With A Finite LatticeDokument4 SeitenAlexey Chilikov, Alexey Zhukov, Alexey Verkhovsky. The Research of Reversible Cellular Automata With A Finite LatticeAlex ZNoch keine Bewertungen
- Cellular Automata With MathematicaDokument9 SeitenCellular Automata With MathematicaDennis La CoteraNoch keine Bewertungen
- 1.1 Nonlinear Model and Nonlinear Phenomena: U U X X T F X U U X X T F XDokument7 Seiten1.1 Nonlinear Model and Nonlinear Phenomena: U U X X T F X U U X X T F XGuilherme Brolin GatoNoch keine Bewertungen
- Chaotic Dynamics of The Fractional Lorenz SystemDokument4 SeitenChaotic Dynamics of The Fractional Lorenz SystemCristian Camilo CerónNoch keine Bewertungen
- Universality CADokument35 SeitenUniversality CAGutemberg AlvesNoch keine Bewertungen
- A Control Lyapunov Function Approach To Robust Stabilization of Nonlinear Systems'Dokument5 SeitenA Control Lyapunov Function Approach To Robust Stabilization of Nonlinear Systems'Saeb AmirAhmadi ChomacharNoch keine Bewertungen
- Feedback Stabilization Via Rational Control Lyapunov FunctionsDokument6 SeitenFeedback Stabilization Via Rational Control Lyapunov FunctionsSaeb AmirAhmadi ChomacharNoch keine Bewertungen
- Logistic Map Circuit PDFDokument8 SeitenLogistic Map Circuit PDFmenguemengueNoch keine Bewertungen
- Numerical Simulation of Unidirectional Chaotic Synchronization of Non-Autonomous Circuit and Its Application For Secure CommunicationDokument13 SeitenNumerical Simulation of Unidirectional Chaotic Synchronization of Non-Autonomous Circuit and Its Application For Secure CommunicationMada Sanjaya WsNoch keine Bewertungen
- Cellular Automata: An Introduction: Gustavo Camelo NetoDokument24 SeitenCellular Automata: An Introduction: Gustavo Camelo NetoGutemberg AlvesNoch keine Bewertungen
- Cellular Automata: Jarkko KariDokument16 SeitenCellular Automata: Jarkko KariallisonarchitectureNoch keine Bewertungen
- Cellular Automata: BY-Rishab Mehta D1703B51 Bt-Mba CseDokument10 SeitenCellular Automata: BY-Rishab Mehta D1703B51 Bt-Mba CseRishab MehtaNoch keine Bewertungen
- Dictyostelium Amoebae Are Free Living Cells With A Remarkable Twist: Under The Stress of StarvationDokument4 SeitenDictyostelium Amoebae Are Free Living Cells With A Remarkable Twist: Under The Stress of StarvationNaresh Prasad SapkotaNoch keine Bewertungen
- Computation at The Edge of Chaos Langton PDFDokument26 SeitenComputation at The Edge of Chaos Langton PDFyu8Noch keine Bewertungen
- Basins of Attraction For Chimera States: A, B C, D D, E, FDokument23 SeitenBasins of Attraction For Chimera States: A, B C, D D, E, Falpha_numericNoch keine Bewertungen
- Cellular Automata in Public Key CryptographyDokument8 SeitenCellular Automata in Public Key CryptographyIJERAS-International Journal of Engineering Research and Applied Science (ISSN: 2349-4522)Noch keine Bewertungen
- Lec 4 Paper - Cellular Neural NetworkDokument13 SeitenLec 4 Paper - Cellular Neural NetworkPraveena AnnaduraiNoch keine Bewertungen
- Cellular Automata Are GenericDokument16 SeitenCellular Automata Are GenericgnugplNoch keine Bewertungen
- SY Lecture015.Cellular AutomataDokument177 SeitenSY Lecture015.Cellular AutomatarizalsscNoch keine Bewertungen
- Lecture 6: Stability of Dynamic Systems Lyapunov's Direct Method 6.1 Notions of StabilityDokument10 SeitenLecture 6: Stability of Dynamic Systems Lyapunov's Direct Method 6.1 Notions of Stabilitytennis1949Noch keine Bewertungen
- The Power of Totalistic Cellular Automata and To - 1988 - Mathematical and CompuDokument3 SeitenThe Power of Totalistic Cellular Automata and To - 1988 - Mathematical and CompudarshanNoch keine Bewertungen
- NASPI March2015 Poster-EPRI-WSU PDFDokument1 SeiteNASPI March2015 Poster-EPRI-WSU PDFInsaf MohamedNoch keine Bewertungen
- A New Chaotic Attractor From General Lorenz System Family and Its Electronic Experimental ImplementationDokument14 SeitenA New Chaotic Attractor From General Lorenz System Family and Its Electronic Experimental Implementationihsan333Noch keine Bewertungen
- Detection of Transiently Chaotic Swings in Power Systems Using Real-Time Phasor MeasurementsDokument8 SeitenDetection of Transiently Chaotic Swings in Power Systems Using Real-Time Phasor MeasurementsChetan MishraNoch keine Bewertungen
- Cellular Automata - 3Dokument41 SeitenCellular Automata - 3Muhanad Al-khalisyNoch keine Bewertungen
- UKF PaperDokument5 SeitenUKF Papertys7524Noch keine Bewertungen
- Microcontroller: CommunicationDokument2 SeitenMicrocontroller: CommunicationAla'a Ddin ShdefatNoch keine Bewertungen
- Mehra 1970Dokument10 SeitenMehra 1970Imane IdrissiNoch keine Bewertungen
- Theory:: Sequence DetetectorDokument2 SeitenTheory:: Sequence DetetectorNarendra AchariNoch keine Bewertungen
- (EXE) Random Walknetwork SDokument7 Seiten(EXE) Random Walknetwork SChristian F. VegaNoch keine Bewertungen
- Theory of Memristive Controllers: Design and Stability Analysis For Linear PlantsDokument9 SeitenTheory of Memristive Controllers: Design and Stability Analysis For Linear PlantsVijyan MahajanNoch keine Bewertungen
- Design of A Patch Antenna With Integration Cellular Automata and Genetic AlgorithmDokument6 SeitenDesign of A Patch Antenna With Integration Cellular Automata and Genetic AlgorithmjozammNoch keine Bewertungen
- BDS Unit 2 - NoRestrictionDokument41 SeitenBDS Unit 2 - NoRestrictionRameshNoch keine Bewertungen
- Cellular Automata Are Idealized Models of Complex SystemsDokument58 SeitenCellular Automata Are Idealized Models of Complex SystemshigginscribdNoch keine Bewertungen
- Universal LearnerDokument9 SeitenUniversal LearnerÓscar Julián Quiroga GarzónNoch keine Bewertungen
- ACS Assignment 1Dokument2 SeitenACS Assignment 1Gadde DeviNoch keine Bewertungen
- Mariusz H. Jakubowski, Ken Steiglitz and Richard K. Squier - When Can Solitons Compute?Dokument23 SeitenMariusz H. Jakubowski, Ken Steiglitz and Richard K. Squier - When Can Solitons Compute?ldkgfdNoch keine Bewertungen
- Linear SuperpositionDokument6 SeitenLinear SuperpositionKatarzyna JankowskaNoch keine Bewertungen
- 0807 2107 PDFDokument61 Seiten0807 2107 PDFAlexandra BarrosoNoch keine Bewertungen
- Automata and CLM Modelling With Distributed Computing: The Constellation Project December 4, 2011Dokument8 SeitenAutomata and CLM Modelling With Distributed Computing: The Constellation Project December 4, 2011Sean DerSchaf ConNoch keine Bewertungen
- mamatAMS1 4 2013 2Dokument11 SeitenmamatAMS1 4 2013 2Mada Sanjaya WsNoch keine Bewertungen
- Adaptive Observer-Based Non-Linear Stochastic System Control With Sliding Mode SchemesDokument10 SeitenAdaptive Observer-Based Non-Linear Stochastic System Control With Sliding Mode Schemes2022 ISSPMNoch keine Bewertungen
- Global Stochastic Synchronization of Chaotic OscillatorsDokument6 SeitenGlobal Stochastic Synchronization of Chaotic OscillatorsBit32Noch keine Bewertungen
- Jerk Equation PDFDokument4 SeitenJerk Equation PDFmenguemengueNoch keine Bewertungen
- 10 0000@ieeexplore Ieee Org@7076162Dokument6 Seiten10 0000@ieeexplore Ieee Org@7076162ghassen marouaniNoch keine Bewertungen
- Sliding Mode Control Based On Observers For Uncertain Nonlinear SystemsDokument6 SeitenSliding Mode Control Based On Observers For Uncertain Nonlinear SystemsWaj DINoch keine Bewertungen
- Lyapunov MITDokument18 SeitenLyapunov MITAnwesha MukhopadhyayNoch keine Bewertungen
- Cellular Neural Networks: Theory: Stability AnalysisDokument16 SeitenCellular Neural Networks: Theory: Stability AnalysisMauricio RamosNoch keine Bewertungen
- Applications of Dynamical SystemsDokument32 SeitenApplications of Dynamical SystemsAl VlearNoch keine Bewertungen
- CMR900Dokument6 SeitenCMR900Time To Use BrainNoch keine Bewertungen
- Sandisk X400 SSD (Solid State Drive) : The World'S First 1Tb, Single-Sided SSDDokument2 SeitenSandisk X400 SSD (Solid State Drive) : The World'S First 1Tb, Single-Sided SSDlechurzNoch keine Bewertungen
- MultiplexersDokument23 SeitenMultiplexersAsim WarisNoch keine Bewertungen
- Trial On CompresorDokument3 SeitenTrial On CompresorA JNoch keine Bewertungen
- RS232STATUSMONDokument1 SeiteRS232STATUSMONashish.boradNoch keine Bewertungen
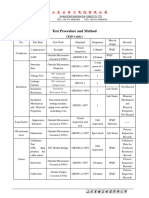
- Test Procedure and MethodDokument1 SeiteTest Procedure and MethodkmiqdNoch keine Bewertungen
- Chapter 7 Series Parallel CircuitsDokument70 SeitenChapter 7 Series Parallel CircuitsAbdullrahman Al-ShammaaNoch keine Bewertungen
- Family Surveyed 2017 - TimberDokument26 SeitenFamily Surveyed 2017 - TimberAlibasher Macalnas0% (1)
- RFQ - Tank DemuckingDokument12 SeitenRFQ - Tank Demuckingmuhamadrafie1975100% (1)
- Heat Loss and Gain CalculationDokument84 SeitenHeat Loss and Gain Calculationafraz_xecNoch keine Bewertungen
- Java Lang OutOfMemoryError Handbook - PlumbrDokument28 SeitenJava Lang OutOfMemoryError Handbook - PlumbrcuonglunNoch keine Bewertungen
- 20 and 21. Requirements For Licensure by Examination Nclex. Revised 06.20.19 1Dokument2 Seiten20 and 21. Requirements For Licensure by Examination Nclex. Revised 06.20.19 1Glennah Marie Avenido RamosNoch keine Bewertungen
- CM29, 03-16-17Dokument3 SeitenCM29, 03-16-17Louie PascuaNoch keine Bewertungen
- SWIFT Case Study Accord EADS 200808Dokument3 SeitenSWIFT Case Study Accord EADS 200808maheshbendigeri5945Noch keine Bewertungen
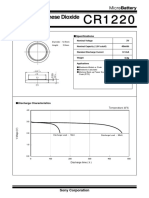
- Sony CR1220 PDFDokument2 SeitenSony CR1220 PDFdiego742000Noch keine Bewertungen
- Capitulo 9 - Flujo de DatosDokument24 SeitenCapitulo 9 - Flujo de DatosOrlando Espinoza ZevallosNoch keine Bewertungen
- ITS Quick Ref GuideDokument6 SeitenITS Quick Ref GuidedhanahbalNoch keine Bewertungen