Das könnte Ihnen auch gefallen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Forced CirculationDokument3 SeitenForced CirculationHyung Sholahuddin AlayNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Problem 5.12Dokument2 SeitenProblem 5.12Hyung Sholahuddin AlayNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- 090Dokument6 Seiten090Abdul Mannan KhanNoch keine Bewertungen
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Title - 16 Words at Max Capitalize The Initial Letter of Each Word Excluding The Articles and PrepositionsDokument2 SeitenTitle - 16 Words at Max Capitalize The Initial Letter of Each Word Excluding The Articles and PrepositionsHyung Sholahuddin AlayNoch keine Bewertungen
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Stripping StudyDokument3 SeitenStripping StudyHyung Sholahuddin AlayNoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Jadwal Akademik 2016-2017 - 1Dokument2 SeitenJadwal Akademik 2016-2017 - 1Hyung Sholahuddin AlayNoch keine Bewertungen
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Molecules: Biological and Nutritional Properties of Palm Oil and Palmitic Acid: Effects On HealthDokument23 SeitenMolecules: Biological and Nutritional Properties of Palm Oil and Palmitic Acid: Effects On HealthHyung Sholahuddin AlayNoch keine Bewertungen
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- FormaldehydeDokument7 SeitenFormaldehydeAsih MiniartiNoch keine Bewertungen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Aspen User Guide 10Dokument936 SeitenAspen User Guide 10tryinghard18Noch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- 090Dokument6 Seiten090Abdul Mannan KhanNoch keine Bewertungen
- Production of Synthesis Gas: Caalysis Today, 18 (1993) 305-324Dokument20 SeitenProduction of Synthesis Gas: Caalysis Today, 18 (1993) 305-324ainmnrhNoch keine Bewertungen
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Ethanol Production From Sago Waste Using Saccharomyces Cerevisiae Vits-M1Dokument10 SeitenEthanol Production From Sago Waste Using Saccharomyces Cerevisiae Vits-M1Hyung Sholahuddin AlayNoch keine Bewertungen
- Chapter 16 - Sulphur: The Frasch Process Uses of Sulphur Allotropes of Sulphur Properties of Sulphur Sulphur DioxideDokument28 SeitenChapter 16 - Sulphur: The Frasch Process Uses of Sulphur Allotropes of Sulphur Properties of Sulphur Sulphur DioxideHyung Sholahuddin AlayNoch keine Bewertungen
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Solution Week 9Dokument6 SeitenSolution Week 9Ariadne ChuaNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Solution Week 9Dokument6 SeitenSolution Week 9Ariadne ChuaNoch keine Bewertungen
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Latihan IELTSDokument1 SeiteLatihan IELTSHyung Sholahuddin AlayNoch keine Bewertungen
- Fundamentals of ElectrochemistryDokument60 SeitenFundamentals of ElectrochemistryHyung Sholahuddin AlayNoch keine Bewertungen
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- BooeDokument5 SeitenBooeHyung Sholahuddin AlayNoch keine Bewertungen
- Water RemovalDokument2 SeitenWater RemovalHyung Sholahuddin AlayNoch keine Bewertungen
- DiffusionToCloudDroplet 1Dokument7 SeitenDiffusionToCloudDroplet 1Hyung Sholahuddin AlayNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Chapter 16 - Sulphur: The Frasch Process Uses of Sulphur Allotropes of Sulphur Properties of Sulphur Sulphur DioxideDokument28 SeitenChapter 16 - Sulphur: The Frasch Process Uses of Sulphur Allotropes of Sulphur Properties of Sulphur Sulphur DioxideHyung Sholahuddin AlayNoch keine Bewertungen
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Asli EditDokument6 SeitenAsli EditHyung Sholahuddin AlayNoch keine Bewertungen
- PR Kimia AnalisisDokument7 SeitenPR Kimia AnalisisHyung Sholahuddin AlayNoch keine Bewertungen
- SEED Guide Section 2 - Seed Saving, Sharing, GrowingDokument9 SeitenSEED Guide Section 2 - Seed Saving, Sharing, GrowingMaggie SkylerNoch keine Bewertungen
- Dalip Kumar Upreti, Pradeep K. Divakar, Vertika Shukla, Rajesh Bajpai (Eds.)-Recent Advances in Lichenology_ Modern Methods and Approaches in Lichen Systematics and Culture Techniques, Volume 2-SpringDokument240 SeitenDalip Kumar Upreti, Pradeep K. Divakar, Vertika Shukla, Rajesh Bajpai (Eds.)-Recent Advances in Lichenology_ Modern Methods and Approaches in Lichen Systematics and Culture Techniques, Volume 2-SpringvitaNoch keine Bewertungen
- COMBINED SCIENCE1 Min PDFDokument100 SeitenCOMBINED SCIENCE1 Min PDFLovemore MalakiNoch keine Bewertungen
- RHH Family Mealtime ToolkitDokument79 SeitenRHH Family Mealtime ToolkittherealhappyhourNoch keine Bewertungen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Monthly Narrative ReportDokument14 SeitenMonthly Narrative ReportDing CadavisNoch keine Bewertungen
- Menu ProjectDokument1 SeiteMenu Projectapi-357256089Noch keine Bewertungen
- 20170918140947SBC 3023 - TBB 2033beditednew4Dokument36 Seiten20170918140947SBC 3023 - TBB 2033beditednew4Leena MuniandyNoch keine Bewertungen
- Biology Lecture Notes: (Stemer'S Guide)Dokument22 SeitenBiology Lecture Notes: (Stemer'S Guide)AbdoNoch keine Bewertungen
- CCI Buyers Guide 2020Dokument47 SeitenCCI Buyers Guide 2020Chris JonesNoch keine Bewertungen
- Tree ParentingDokument10 SeitenTree ParentingSophia Anne MonillasNoch keine Bewertungen
- Agrobacterium RhizogenesDokument37 SeitenAgrobacterium Rhizogeneskrishna16100% (2)
- Fu Talisman For Physic HealthDokument6 SeitenFu Talisman For Physic HealthmesiaNoch keine Bewertungen
- Program Book HSS & The 6th ISSH 2016 PDFDokument141 SeitenProgram Book HSS & The 6th ISSH 2016 PDFAkaz PujoNoch keine Bewertungen
- Sol KadhiDokument1 SeiteSol KadhiSourabh DholeNoch keine Bewertungen
- NCERT Book Class 3 Enviromental Studies Looking Around Chapter 2 The Plant Fairy PDFDokument9 SeitenNCERT Book Class 3 Enviromental Studies Looking Around Chapter 2 The Plant Fairy PDFkarthiyuvenNoch keine Bewertungen
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Aquarium PDFDokument34 SeitenAquarium PDFsksingl350Noch keine Bewertungen
- Pengaruh Pupuk Organik Terhadap Pertumbuhan Dan Hasil Tanaman Tomat Pada Tanah Aluvial OlehDokument7 SeitenPengaruh Pupuk Organik Terhadap Pertumbuhan Dan Hasil Tanaman Tomat Pada Tanah Aluvial OlehbaitNoch keine Bewertungen
- Best Practice For Pilot Plant LayoutDokument10 SeitenBest Practice For Pilot Plant LayoutRashesh ShahNoch keine Bewertungen
- KS TRB LC Read SumDokument7 SeitenKS TRB LC Read Sumgabriela Fernandez MoiNoch keine Bewertungen
- Black Spine Book 2Dokument98 SeitenBlack Spine Book 2Raymond MicheauNoch keine Bewertungen
- Hybrid Seed Production in CottonDokument27 SeitenHybrid Seed Production in Cottonnarayanan100% (1)
- Flavonols and Phenolic Acids in Berries and Berry Products PDFDokument93 SeitenFlavonols and Phenolic Acids in Berries and Berry Products PDFGopalaKrishnan SivaramanNoch keine Bewertungen
- Grade 6 DLL ESP 6 Q1 Week 3abcDokument7 SeitenGrade 6 DLL ESP 6 Q1 Week 3abcGeizel ReubalNoch keine Bewertungen
- Seminar Functional DrinksDokument22 SeitenSeminar Functional DrinksMoshmi MazumdarNoch keine Bewertungen
- Modern Biology. Chapter Tests With Answer Key General and AdvancedDokument250 SeitenModern Biology. Chapter Tests With Answer Key General and AdvancedWllatakam100% (7)
- AbbreviationsDokument6 SeitenAbbreviationskalidas_295Noch keine Bewertungen
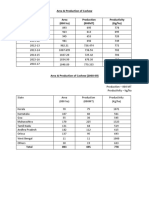
- Area & Production of Cashew Area (000 Ha) Production (000MT) Productivity (Kg/ha)Dokument6 SeitenArea & Production of Cashew Area (000 Ha) Production (000MT) Productivity (Kg/ha)DivyeshNoch keine Bewertungen
- 80 Square Feet SHTF Medicinal GardenDokument42 Seiten80 Square Feet SHTF Medicinal Gardenemily turner100% (3)
- Pokémon Go IV Calculator - Game DataDokument6 SeitenPokémon Go IV Calculator - Game DataMax KeltonNoch keine Bewertungen
- Pure Bio CH 7 Textbook Answers PDFDokument3 SeitenPure Bio CH 7 Textbook Answers PDFlee50% (8)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisVon EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisBewertung: 3.5 von 5 Sternen3.5/5 (2)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceVon EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceBewertung: 4.5 von 5 Sternen4.5/5 (516)
- Tales from Both Sides of the Brain: A Life in NeuroscienceVon EverandTales from Both Sides of the Brain: A Life in NeuroscienceBewertung: 3 von 5 Sternen3/5 (18)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Von EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Bewertung: 4 von 5 Sternen4/5 (378)
- Gut: the new and revised Sunday Times bestsellerVon EverandGut: the new and revised Sunday Times bestsellerBewertung: 4 von 5 Sternen4/5 (392)