Das könnte Ihnen auch gefallen
- Alteraciones genéticas infantilesDokument36 SeitenAlteraciones genéticas infantilesItzel GómezNoch keine Bewertungen
- Compilación Artículos de Ernest Rossi - ADokument212 SeitenCompilación Artículos de Ernest Rossi - ADavid Alejandro Villeda100% (2)
- Sesión 04 Tema, Subtema - EtcDokument6 SeitenSesión 04 Tema, Subtema - EtcJuana100% (4)
- Partes de Una MaquinaDokument16 SeitenPartes de Una MaquinaEdsonPagani100% (1)
- Autismo y MicrobiotaDokument14 SeitenAutismo y MicrobiotaGloria ArgotiNoch keine Bewertungen
- Plantas Comestibles de La Región de Alvarado, VeracruzDokument31 SeitenPlantas Comestibles de La Región de Alvarado, VeracruzWera Uscanga100% (4)
- La Trata de Personas en El PerúDokument5 SeitenLa Trata de Personas en El PerúEdsonPaganiNoch keine Bewertungen
- El Científico Robert Brawn Fue Quien Descubrió El Núcleo de La Célula en El Año 1665. Nota CientíficaDokument3 SeitenEl Científico Robert Brawn Fue Quien Descubrió El Núcleo de La Célula en El Año 1665. Nota CientíficaIvan Tacza SantanaNoch keine Bewertungen
- Infecciones urinarias: causas, síntomas y tratamientoDokument34 SeitenInfecciones urinarias: causas, síntomas y tratamientoOlimpia GutierrezNoch keine Bewertungen
- Clasificación de Los Seres VivosDokument2 SeitenClasificación de Los Seres VivosArthuriuzMRamosNoch keine Bewertungen
- 3 Regimen TributarioDokument2 Seiten3 Regimen TributarioEdsonPaganiNoch keine Bewertungen
- 3 Regimen TributarioDokument6 Seiten3 Regimen TributarioLizbeth Cordova YucraNoch keine Bewertungen
- Ciclo de Vida 2Dokument12 SeitenCiclo de Vida 2EdsonPaganiNoch keine Bewertungen
- Musculo EsqueleticoDokument1 SeiteMusculo EsqueleticoEdsonPaganiNoch keine Bewertungen
- Juego en El Desarrollo Del NiñoDokument8 SeitenJuego en El Desarrollo Del NiñoEdsonPaganiNoch keine Bewertungen
- Sesion JunioDokument33 SeitenSesion JunioEdsonPaganiNoch keine Bewertungen
- 5 Platos Tipicos de La Region SudamericanaDokument14 Seiten5 Platos Tipicos de La Region SudamericanaEdsonPaganiNoch keine Bewertungen
- Ficha Inscripción Maestría EPGDokument1 SeiteFicha Inscripción Maestría EPGEdsonPaganiNoch keine Bewertungen
- El CalostroDokument3 SeitenEl CalostroEdsonPaganiNoch keine Bewertungen
- Que Es El Programa de CUNAMASDokument9 SeitenQue Es El Programa de CUNAMASEdsonPaganiNoch keine Bewertungen
- InformeDokument1 SeiteInformeEdsonPaganiNoch keine Bewertungen
- AnemiaDokument1 SeiteAnemiaEdsonPaganiNoch keine Bewertungen
- AMORTIGUADORESDokument13 SeitenAMORTIGUADORESEdsonPaganiNoch keine Bewertungen
- AnemiaDokument2 SeitenAnemiaEdsonPaganiNoch keine Bewertungen
- Pollo Frito Con ArrozDokument4 SeitenPollo Frito Con ArrozEdsonPaganiNoch keine Bewertungen
- Pastos y ForrajesDokument35 SeitenPastos y ForrajesEdsonPagani100% (1)
- Resumen de La Ley General de SociedadesDokument7 SeitenResumen de La Ley General de SociedadesEdsonPaganiNoch keine Bewertungen
- SolicitudDokument1 SeiteSolicitudEdsonPaganiNoch keine Bewertungen
- Bambu PomaDokument53 SeitenBambu PomaEdsonPaganiNoch keine Bewertungen
- Arroz Con Pollo FritoDokument5 SeitenArroz Con Pollo FritoEdsonPaganiNoch keine Bewertungen
- Partes de Una MaquinaDokument1 SeitePartes de Una MaquinaEdsonPaganiNoch keine Bewertungen
- Historia y origen del arroz con leche peruanoDokument5 SeitenHistoria y origen del arroz con leche peruanoEdsonPaganiNoch keine Bewertungen
- Elaboración de Una Bebida Antioxidante Proyecto Crea y EmprendeDokument6 SeitenElaboración de Una Bebida Antioxidante Proyecto Crea y EmprendeEdsonPaganiNoch keine Bewertungen
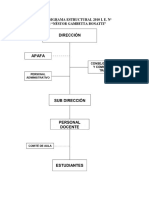
- Organigrama Estructural 2010 IDokument13 SeitenOrganigrama Estructural 2010 IEdsonPaganiNoch keine Bewertungen
- Contrato de Trabajo de Villa RicaDokument1 SeiteContrato de Trabajo de Villa RicaEdsonPaganiNoch keine Bewertungen
- Informe #02-2018 MAYO ISAAC - OXAPDokument12 SeitenInforme #02-2018 MAYO ISAAC - OXAPEdsonPaganiNoch keine Bewertungen
- Ficha N 01Dokument2 SeitenFicha N 01EdsonPaganiNoch keine Bewertungen
- Manual de Geriatría y Gerontología PUCPDokument130 SeitenManual de Geriatría y Gerontología PUCPmavcc100% (1)
- Pedic Ulos IsDokument18 SeitenPedic Ulos IsRosario Medalit Quezada PacoraNoch keine Bewertungen
- Quinolonas PDFDokument7 SeitenQuinolonas PDFDiego PMNoch keine Bewertungen
- Gel AntibacterialDokument32 SeitenGel AntibacterialEric Franco Pimentel57% (7)
- Micro Informe 2Dokument33 SeitenMicro Informe 2Jimy De La Cruz Carhuallanqui100% (1)
- Genetica Grupo 2Dokument50 SeitenGenetica Grupo 2Cindy RomeroNoch keine Bewertungen
- MtecsDokument17 SeitenMtecssandraNoch keine Bewertungen
- Estimado Comprensivo Del Paciente Con Condición CrónicaDokument16 SeitenEstimado Comprensivo Del Paciente Con Condición CrónicaLeyenda93Noch keine Bewertungen
- 13 7 Retraso LectorDokument2 Seiten13 7 Retraso LectorViviana VegaNoch keine Bewertungen
- El Metodo ClinicoDokument11 SeitenEl Metodo ClinicoJc FigueroaNoch keine Bewertungen
- CelulaDokument71 SeitenCelulajackerondonNoch keine Bewertungen
- Vitamina KDokument18 SeitenVitamina Kdeisy_jasmin17Noch keine Bewertungen
- El ChileDokument3 SeitenEl ChileOsmar Soto50% (6)
- Estructura y Función de Nucleótidos 1Dokument11 SeitenEstructura y Función de Nucleótidos 1Mauricio Pulido JiménezNoch keine Bewertungen
- Tema 3. Asepsia PDFDokument17 SeitenTema 3. Asepsia PDFRocío Martínez MartínezNoch keine Bewertungen
- Inmunologia CompletoDokument59 SeitenInmunologia CompletoSebastián AgudeloNoch keine Bewertungen
- Abestosis y Mesotelioma Pleural MalignoDokument13 SeitenAbestosis y Mesotelioma Pleural MalignoAlonso LopezNoch keine Bewertungen
- Ciclo CelularDokument4 SeitenCiclo CelularMilagros Delfina ColombiniNoch keine Bewertungen
- Principales Enfermedades en AdolescentesDokument6 SeitenPrincipales Enfermedades en AdolescentesObitoHuchihaNoch keine Bewertungen
- Qué Es Kotsuban Yumeiho - y Su Teoria de Fundamento en Desnivel de Las CaderasDokument3 SeitenQué Es Kotsuban Yumeiho - y Su Teoria de Fundamento en Desnivel de Las CaderasEduardo Baquerizo WongNoch keine Bewertungen
- Formulas Enterales Adulto y PediatriaDokument23 SeitenFormulas Enterales Adulto y Pediatriacarolina100% (1)
- Cáncer de Mama y CerveixDokument3 SeitenCáncer de Mama y CerveixMax Herrera VargasNoch keine Bewertungen
- Extracción Del AdnDokument10 SeitenExtracción Del AdnAngeliño Oña MamaniNoch keine Bewertungen