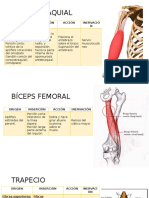
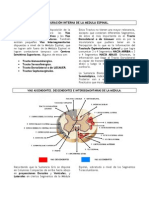
Das könnte Ihnen auch gefallen
- Pruebas Funcional Muscular - Daniels-6 Edic - 220315 - 173224Dokument425 SeitenPruebas Funcional Muscular - Daniels-6 Edic - 220315 - 173224Cris Vázquez100% (3)
- Triptico La Adolecencia Esamju PDFDokument2 SeitenTriptico La Adolecencia Esamju PDFSteven Berrios100% (4)
- Dermatomas del cuerpo humanoDokument1 SeiteDermatomas del cuerpo humanoMonse100% (5)
- Mi Dieta Anticancer PDFDokument15 SeitenMi Dieta Anticancer PDFLady_JacNoch keine Bewertungen
- Fascículo Grácil y CuneiformeDokument3 SeitenFascículo Grácil y CuneiformeCarolina Urrutia100% (3)
- Pruebas funcionales hombro y codoDokument5 SeitenPruebas funcionales hombro y codoNataniel FloresNoch keine Bewertungen
- Test KendallDokument7 SeitenTest KendallIrel RivadeneiraNoch keine Bewertungen
- Dana Marie Bell Serie Manada Poconos I Encontrando El PerdonDokument120 SeitenDana Marie Bell Serie Manada Poconos I Encontrando El PerdonLorena Cruz Hernandez Muñiz100% (6)
- Dermatomas y MiotomasDokument29 SeitenDermatomas y MiotomasMaria Celeste67% (3)
- Dermatomas y Miotomas ActualizadoDokument3 SeitenDermatomas y Miotomas ActualizadoRaymi Luis Ramirez100% (2)
- Anatomia Del Plexo Lumbar y LumbosacroDokument2 SeitenAnatomia Del Plexo Lumbar y LumbosacroRuth Pacheco0% (1)
- Epicondilitis PDFDokument6 SeitenEpicondilitis PDFTiitiimuaNoch keine Bewertungen
- Tratamiento Fisioterapéutico en Fracturas de Radio y CubitoDokument28 SeitenTratamiento Fisioterapéutico en Fracturas de Radio y CubitoLuis Bazán Pino100% (1)
- Vias ExtrapiramidalesDokument10 SeitenVias ExtrapiramidalesElizabeth Auccapiña GomezNoch keine Bewertungen
- Paralisis de BellDokument1 SeiteParalisis de BellNury Ramirez100% (1)
- Exploración de Dermatomas y Miotomas, Esclerotoma y ViscerotomaDokument11 SeitenExploración de Dermatomas y Miotomas, Esclerotoma y ViscerotomaMonserrat Cruz GómezNoch keine Bewertungen
- Via Espinotalamica Lateral y AnteriorDokument1 SeiteVia Espinotalamica Lateral y AnteriorPaula Gaviria50% (2)
- Prueba de AdamsDokument12 SeitenPrueba de Adamstonni marchena100% (1)
- Método de KaltenbornDokument15 SeitenMétodo de KaltenbornAlexis BernillaNoch keine Bewertungen
- Evaluación Respiratoria Completa enDokument75 SeitenEvaluación Respiratoria Completa enTiitiimuaNoch keine Bewertungen
- Puntos MotoresDokument1 SeitePuntos MotoresLucia Petrone83% (6)
- Las Vías SensitivasDokument5 SeitenLas Vías SensitivasLeodan Garcia YbañezNoch keine Bewertungen
- Cancer - Herencia y Ambiente - Cristina CortinasDokument50 SeitenCancer - Herencia y Ambiente - Cristina Cortinascarabe100% (3)
- Cuadro Comparativo de Técnicas de Tratamiento en Fisioterapia Pediatrica.Dokument16 SeitenCuadro Comparativo de Técnicas de Tratamiento en Fisioterapia Pediatrica.Madelene Barrios100% (3)
- Síndrome de Neurona Motora 2Dokument26 SeitenSíndrome de Neurona Motora 2Mara Karyme SuasteNoch keine Bewertungen
- Ejercicios de RisserDokument1 SeiteEjercicios de RisserDiana CepedaNoch keine Bewertungen
- Aislamiento de HongosDokument28 SeitenAislamiento de HongosNelson Conislla CastilloNoch keine Bewertungen
- PAP en Niños y AdolescentesDokument64 SeitenPAP en Niños y Adolescentesfredis rodriguweszNoch keine Bewertungen
- Sdra: EpocDokument59 SeitenSdra: EpocTiitiimuaNoch keine Bewertungen
- Sdra: EpocDokument59 SeitenSdra: EpocTiitiimuaNoch keine Bewertungen
- Laminas de RexedDokument2 SeitenLaminas de Rexedtunaantes83% (6)
- Introducción A La Evaluación Kinésica y EntrevistaDokument47 SeitenIntroducción A La Evaluación Kinésica y EntrevistaNicole Vargas Marín100% (1)
- Anatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Von EverandAnatomia y Fisiología del Sistema Nervioso II: Principios Elementales del Sistema Nervioso, #2Noch keine Bewertungen
- Reflejos OsteotendinososDokument18 SeitenReflejos OsteotendinososSofía SalasNoch keine Bewertungen
- Guía cumplimiento protocolo vigilancia plaguicidasDokument38 SeitenGuía cumplimiento protocolo vigilancia plaguicidasangelicaalave2012Noch keine Bewertungen
- Clasificacion de La Sensibilidad Superficial y ProfundaDokument1 SeiteClasificacion de La Sensibilidad Superficial y ProfundaVictoria Albarran100% (2)
- Línea Del Tiempo Historia de La Terapia OcupacionalDokument4 SeitenLínea Del Tiempo Historia de La Terapia OcupacionalShannon Rodriguez100% (3)
- Vias Ascendentes y Descendentes de La Medula EspinalDokument6 SeitenVias Ascendentes y Descendentes de La Medula EspinalSarai LopezNoch keine Bewertungen
- Pares CranealesDokument3 SeitenPares CranealesRocio Cardenas100% (3)
- Bíceps BraquialDokument6 SeitenBíceps BraquialBrenda Gress Viveros0% (1)
- Electroterapia en El Paciente QuemadoDokument14 SeitenElectroterapia en El Paciente QuemadoMicaela33% (3)
- Cap. 56 Control de La Función Motora Por La Corteza y El Tronco Del EncéfaloDokument5 SeitenCap. 56 Control de La Función Motora Por La Corteza y El Tronco Del EncéfaloJocelyne Benito100% (3)
- Tipos de Marchas PatológicasDokument22 SeitenTipos de Marchas PatológicasFatima SvNoch keine Bewertungen
- Part 3 CONFIGURACIÓN INTERNA DE LA MEDULA ESPINALDokument12 SeitenPart 3 CONFIGURACIÓN INTERNA DE LA MEDULA ESPINALRaymi Luis Ramirez100% (3)
- Control PosturalDokument72 SeitenControl PosturalZandraTC100% (1)
- Tablas GoniometriaDokument4 SeitenTablas GoniometriaCynthia CamposNoch keine Bewertungen
- Codo Muneca y ManoDokument42 SeitenCodo Muneca y ManoTiitiimuaNoch keine Bewertungen
- Caso Clínico TerryDokument28 SeitenCaso Clínico TerryLizeth Andrea IguadNoch keine Bewertungen
- 11 Principios Basicos de FNPDokument10 Seiten11 Principios Basicos de FNPAna Verónica De león Paiz100% (1)
- Corteza motora: estructura y funciones principalesDokument17 SeitenCorteza motora: estructura y funciones principalesBryan JuarezNoch keine Bewertungen
- T 16 Enfermedades ITS ExponerDokument36 SeitenT 16 Enfermedades ITS ExponerANDREA BERTHA VARGAS MERIDANoch keine Bewertungen
- ATETOSISDokument11 SeitenATETOSISsofia67% (3)
- Método de Intervencion en Fisioterapia (Mif)Dokument16 SeitenMétodo de Intervencion en Fisioterapia (Mif)linkender100% (1)
- Infecciones felinas: Leucemia y VIH felinoDokument31 SeitenInfecciones felinas: Leucemia y VIH felinoLaura TorresNoch keine Bewertungen
- Funciones del sistema extrapiramidal y sus tractosDokument2 SeitenFunciones del sistema extrapiramidal y sus tractosDanner BacaNoch keine Bewertungen
- Via EspinocerebeloDokument4 SeitenVia EspinocerebeloElisa CamposNoch keine Bewertungen
- Casos ClínicosDokument17 SeitenCasos ClínicosmarcelaNoch keine Bewertungen
- Tecnicas SemiologicasDokument44 SeitenTecnicas SemiologicasRevorg zeñaY adecam70% (10)
- Método Rood PDFDokument2 SeitenMétodo Rood PDFLeo_Ram98100% (1)
- Organización, Estructura y Funciones de Las Meninges y Del Líquido CefalorraquídeoDokument4 SeitenOrganización, Estructura y Funciones de Las Meninges y Del Líquido CefalorraquídeoDiego Bernal100% (1)
- Modelo Cinesiologico, Patocinesiologico, CinesipatologicoDokument4 SeitenModelo Cinesiologico, Patocinesiologico, CinesipatologicoANGIE PESEROSNoch keine Bewertungen
- MECANOTERAPIA ExpoDokument31 SeitenMECANOTERAPIA ExpoJohanna Galvis100% (1)
- Constantes y Variantes de La MarchaDokument7 SeitenConstantes y Variantes de La MarchaBerthalila Madrigal100% (1)
- Síndrome Del Cono Medular - Ventura Chuquiano Sofia VannesaDokument7 SeitenSíndrome Del Cono Medular - Ventura Chuquiano Sofia VannesaLucero Malarín100% (1)
- Tractos EspinocerebelososDokument2 SeitenTractos EspinocerebelososAntonellaGodinezSalazarNoch keine Bewertungen
- Exploración Del Viii Par CranealDokument4 SeitenExploración Del Viii Par CranealJesus NavarroNoch keine Bewertungen
- Anatomía Interna Del MesencéfaloDokument34 SeitenAnatomía Interna Del MesencéfaloPaolaLeos100% (9)
- Preguntas Ganglios BasalesDokument7 SeitenPreguntas Ganglios BasalesMichelle Cartín TristánNoch keine Bewertungen
- Escala de fuerza muscular DanielsDokument3 SeitenEscala de fuerza muscular DanielsMauroPogliacomiNoch keine Bewertungen
- Teoria de La Liberación de EndorfinasDokument1 SeiteTeoria de La Liberación de EndorfinasPablo Pallante Astudillo50% (4)
- SinergiasDokument3 SeitenSinergiasCarlos Alvarado0% (1)
- Escala de LovettDokument2 SeitenEscala de LovettYeiny Solano100% (1)
- Istema Piramidal y ExtrapiramidalDokument12 SeitenIstema Piramidal y ExtrapiramidalRenee CollinsNoch keine Bewertungen
- 6-Via Piramidal y ExtrapiramidalDokument31 Seiten6-Via Piramidal y ExtrapiramidalMiriam MendozaNoch keine Bewertungen
- Sistema Extra Piramidal Informe Casi Listo 1Dokument13 SeitenSistema Extra Piramidal Informe Casi Listo 1PalomitadrNoch keine Bewertungen
- Abordaje FT en Cirugía Cardiaca BYPASSDokument56 SeitenAbordaje FT en Cirugía Cardiaca BYPASSTiitiimuaNoch keine Bewertungen
- Patologia Pares 1Dokument13 SeitenPatologia Pares 1Alan SaucedoNoch keine Bewertungen
- 2013 PDFDokument17 Seiten2013 PDFJuan Camilo MosqueraNoch keine Bewertungen
- Evaluacion PosturaDokument3 SeitenEvaluacion PosturaTiitiimuaNoch keine Bewertungen
- Patologia Pares 1Dokument13 SeitenPatologia Pares 1Alan SaucedoNoch keine Bewertungen
- Biomecánica Del Síndrome de Latigazo Cervical y Su Analogía Osteopática PDFDokument9 SeitenBiomecánica Del Síndrome de Latigazo Cervical y Su Analogía Osteopática PDFTiitiimuaNoch keine Bewertungen
- Ventilación mecánicaDokument11 SeitenVentilación mecánicaPablo Andres0% (1)
- 2011Dokument16 Seiten2011emersonpetikNoch keine Bewertungen
- Terapia NebulizadaDokument11 SeitenTerapia NebulizadaTiitiimuaNoch keine Bewertungen
- Traumatologia CaderaDokument10 SeitenTraumatologia CaderaTiitiimuaNoch keine Bewertungen
- MarchaDokument9 SeitenMarchaMiguel Angel Tipiani MallmaNoch keine Bewertungen
- VM DesteteDokument18 SeitenVM DestetegamabeNoch keine Bewertungen
- Practica #3 Auscultaciçon PulmonarDokument41 SeitenPractica #3 Auscultaciçon PulmonarHarold IbrahimNoch keine Bewertungen
- Protocolo Shock AnafilacticoDokument15 SeitenProtocolo Shock AnafilacticoDalia GMNoch keine Bewertungen
- Control Motor y Mov. NormalDokument21 SeitenControl Motor y Mov. NormalTiitiimuaNoch keine Bewertungen
- Entrenamiento Insuficiencia CardiacaDokument9 SeitenEntrenamiento Insuficiencia CardiacaTiitiimuaNoch keine Bewertungen
- Cuadro Ruidos RespiratoriosDokument2 SeitenCuadro Ruidos RespiratoriosTiitiimuaNoch keine Bewertungen
- "Véndele A La Mente y No A La Gente".: Wendy Santamaría SPT 132-35Dokument9 Seiten"Véndele A La Mente y No A La Gente".: Wendy Santamaría SPT 132-35TiitiimuaNoch keine Bewertungen
- Conceptos TERAPIA MANUAL BásicasDokument15 SeitenConceptos TERAPIA MANUAL BásicasTiitiimuaNoch keine Bewertungen
- DisneaDokument3 SeitenDisneaTiitiimuaNoch keine Bewertungen
- Articulo 2.en - EsDokument6 SeitenArticulo 2.en - EsTiitiimuaNoch keine Bewertungen
- PC EspásticaDokument5 SeitenPC EspásticaTiitiimuaNoch keine Bewertungen
- EquimosisDokument9 SeitenEquimosisValeria Bernardo Arellano100% (1)
- 039 - 08uhdaod SfcdoivfisoDokument16 Seiten039 - 08uhdaod SfcdoivfisoFilipe MaqueiroNoch keine Bewertungen
- Iso23875 PeruDokument10 SeitenIso23875 PeruJOSE LUIS ESPINOZA AVENDAÑONoch keine Bewertungen
- Cadena EpidemiologicaDokument27 SeitenCadena EpidemiologicaLuiizito' Urve'Noch keine Bewertungen
- Norma Mexicana TVCDokument13 SeitenNorma Mexicana TVCVictor M MoralesNoch keine Bewertungen
- Curso Mha InduccionDokument66 SeitenCurso Mha InduccionCarlos MonrealNoch keine Bewertungen
- Cómo Descongestionar Los Senos ParanasalesDokument6 SeitenCómo Descongestionar Los Senos ParanasalesMario Piña RomeroNoch keine Bewertungen
- Situacion SocialDokument100 SeitenSituacion SocialcarlospesantzNoch keine Bewertungen
- Analogías y conceptos clave para el aprendizajeDokument4 SeitenAnalogías y conceptos clave para el aprendizajeyeseniaNoch keine Bewertungen
- VestibularDokument89 SeitenVestibularBárbara González CanteroNoch keine Bewertungen
- Órtesis y Trauma AcumulativoDokument4 SeitenÓrtesis y Trauma AcumulativoCecilia Margarita Vera CastilloNoch keine Bewertungen
- Caso Clínico 4Dokument10 SeitenCaso Clínico 4Viridiana GonzalezNoch keine Bewertungen
- Epi TopoDokument3 SeitenEpi TopoGustavo SepulvedaNoch keine Bewertungen
- LeucemiasDokument3 SeitenLeucemiasGabriela DiazNoch keine Bewertungen
- Protocolo HeladeriaDokument5 SeitenProtocolo HeladeriafranciscoNoch keine Bewertungen
- Fracaso Osteosintesis PDFDokument6 SeitenFracaso Osteosintesis PDFDikita ZamaNoch keine Bewertungen
- Casos clinicos herpes simplexDokument16 SeitenCasos clinicos herpes simplexJean FloruNoch keine Bewertungen
- LEUCEMIASDokument16 SeitenLEUCEMIASAllan NovaesNoch keine Bewertungen
- Generalidades de FarmacodinamiaDokument32 SeitenGeneralidades de FarmacodinamiaMitch Peraza100% (2)
- Miel de AbejaDokument6 SeitenMiel de Abejamaryuri arizaNoch keine Bewertungen
- Informe de La VaricelaDokument4 SeitenInforme de La VaricelaJose RoseroNoch keine Bewertungen
- Manual de Plantas Medicinales Guatemala JDMDokument107 SeitenManual de Plantas Medicinales Guatemala JDMJuana Flores CNoch keine Bewertungen