Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Neuropsych Lec 5 Mental Status ExamDokument2 SeitenNeuropsych Lec 5 Mental Status ExamnkivcNoch keine Bewertungen
- Oral LichenoidDokument12 SeitenOral LichenoiddjianNoch keine Bewertungen
- Kuliah Tumor Rongga MulutDokument36 SeitenKuliah Tumor Rongga MulutIceBearNoch keine Bewertungen
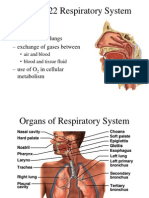
- 22 Respiratory SystemDokument99 Seiten22 Respiratory SystemvanderphysNoch keine Bewertungen
- Hematopoiesis (Occurs in The Liver) Is Considered As The Counterpart in Fetal DevelopmentDokument17 SeitenHematopoiesis (Occurs in The Liver) Is Considered As The Counterpart in Fetal DevelopmentEG Ongteco Gonzaga OngtecoNoch keine Bewertungen
- Nelson Lab ValuesDokument10 SeitenNelson Lab ValuesisahNoch keine Bewertungen
- NSG 126 Serologic Studies (Part 5-8)Dokument11 SeitenNSG 126 Serologic Studies (Part 5-8)Angelica Charisse BuliganNoch keine Bewertungen
- HCC Surveillance in An Era of BiomarkersDokument48 SeitenHCC Surveillance in An Era of BiomarkersRobert G. Gish, MDNoch keine Bewertungen
- Gene TherapyDokument36 SeitenGene TherapySukesh Potla100% (2)
- Pharmacokinetics and Drug Metabolism in The Elderly: Ulrich KlotzDokument10 SeitenPharmacokinetics and Drug Metabolism in The Elderly: Ulrich KlotzMonica LucaciuNoch keine Bewertungen
- Cytokines - IntroductionDokument2 SeitenCytokines - IntroductionTra gicNoch keine Bewertungen
- A. I. in HealthcareDokument14 SeitenA. I. in HealthcaremjNoch keine Bewertungen
- DNA Profiling Gizmo Student Exploration SheetDokument5 SeitenDNA Profiling Gizmo Student Exploration SheetCarter BonesNoch keine Bewertungen
- NWG 2009Dokument316 SeitenNWG 2009globidiNoch keine Bewertungen
- Mengeles ChildrenDokument3 SeitenMengeles Childrenapi-309154321Noch keine Bewertungen
- 2021 ASO Biology Exam Part 1Dokument20 Seiten2021 ASO Biology Exam Part 1Joshua DeathNoch keine Bewertungen
- Argumentative Essay 1Dokument5 SeitenArgumentative Essay 1api-451529335Noch keine Bewertungen
- PredicineATLAS WhitepaperDokument8 SeitenPredicineATLAS WhitepapersagarkarvandeNoch keine Bewertungen
- Komplemen Dan SitokinDokument38 SeitenKomplemen Dan SitokinBantuinAku KakNoch keine Bewertungen
- OriginalDokument24 SeitenOriginalHenrique FerreiraNoch keine Bewertungen
- JC-1 Dye For Mitochondrial Membrane Potential - Thermo Fisher ScientificDokument4 SeitenJC-1 Dye For Mitochondrial Membrane Potential - Thermo Fisher Scientificankitsaneja1Noch keine Bewertungen
- Cerebellar LesionsDokument3 SeitenCerebellar LesionsMary100% (1)
- Surgery Papillary Thyroid CADokument15 SeitenSurgery Papillary Thyroid CAMelissa LabadorNoch keine Bewertungen
- Accurasi Perhitungan PlateletDokument68 SeitenAccurasi Perhitungan PlateletMaria Yosefina HeraNoch keine Bewertungen
- The Benefits of Alpha Lipoic Acid: A Potent AntioxidantDokument56 SeitenThe Benefits of Alpha Lipoic Acid: A Potent AntioxidantAisya Amalia MuslimaNoch keine Bewertungen
- Test Bank For Microbiology An Introduction 13th Edition Gerard J Tortora Berdell R Funke Christine L Case Derek Weber Warner BairDokument35 SeitenTest Bank For Microbiology An Introduction 13th Edition Gerard J Tortora Berdell R Funke Christine L Case Derek Weber Warner Bairshriekacericg31u3100% (37)
- Part 25: Muscle Chapter 216: The Muscular DystrophiesDokument73 SeitenPart 25: Muscle Chapter 216: The Muscular DystrophiesVane100% (1)
- Research PaperDokument28 SeitenResearch PapertaniamanansalabautistaNoch keine Bewertungen
- Bio Control LectureDokument8 SeitenBio Control Lecturedars23Noch keine Bewertungen