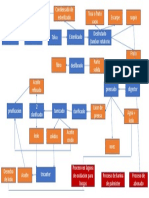
Das könnte Ihnen auch gefallen
- Las agrociencias como soporte a una producción agropecuaria sostenible: Visión ecosistémicaVon EverandLas agrociencias como soporte a una producción agropecuaria sostenible: Visión ecosistémicaNoch keine Bewertungen
- Alternativas para el manejo de residuos sólidos y su integración en el montaje de una huerta agroecológicaVon EverandAlternativas para el manejo de residuos sólidos y su integración en el montaje de una huerta agroecológicaNoch keine Bewertungen
- Curso Vidrio y Platico QuebradizoDokument109 SeitenCurso Vidrio y Platico QuebradizoLuis Eduardo Gomez Guevara100% (3)
- Biorremediacion in SituDokument21 SeitenBiorremediacion in SituJavier Saldarriaga Noé100% (1)
- Los Seis Principios Administrativos de Agustín Reyes PonceDokument10 SeitenLos Seis Principios Administrativos de Agustín Reyes PonceElizabeth Kohatsu0% (2)
- Bituminosos MaterialesDokument41 SeitenBituminosos MaterialesClaudia Yucra SullcaNoch keine Bewertungen
- Experto en gestión medioambientalVon EverandExperto en gestión medioambientalBewertung: 5 von 5 Sternen5/5 (1)
- Biorremediación y FitorremediaciónDokument39 SeitenBiorremediación y FitorremediaciónJuli Jía PCNoch keine Bewertungen
- Andrea Palladio, arquitecto clave del RenacimientoDokument5 SeitenAndrea Palladio, arquitecto clave del RenacimientoDarianna Jimenez100% (1)
- La Pistola HK USP Compact PDFDokument40 SeitenLa Pistola HK USP Compact PDFcholowins100% (1)
- Manual DialuxDokument371 SeitenManual DialuxAngel Fernandes Catire100% (4)
- INET - Nada Es para Siempre Química de La Degradación de Los Materiale PDFDokument181 SeitenINET - Nada Es para Siempre Química de La Degradación de Los Materiale PDFcocolongorro100% (1)
- ITIL V3 Manual CompletoDokument6 SeitenITIL V3 Manual CompletoAlinaAvilesNoch keine Bewertungen
- Ciencias ecologicas. 1983-2013: Treinta años de investigaciones chilenasVon EverandCiencias ecologicas. 1983-2013: Treinta años de investigaciones chilenasNoch keine Bewertungen
- Plan de Contigencia (Ejemplo)Dokument22 SeitenPlan de Contigencia (Ejemplo)IST Francisco Ricci88% (17)
- Biorremediación de AguaDokument19 SeitenBiorremediación de AguaPhamelitha EdquènNoch keine Bewertungen
- Biorremediación de Contaminación Por Mercurio Debido A La Minería Informal en Madre de DiosDokument45 SeitenBiorremediación de Contaminación Por Mercurio Debido A La Minería Informal en Madre de DiosEder Jaime Mamani Pimentel100% (1)
- TrabajoDokument18 SeitenTrabajoSaul Huillcas SNoch keine Bewertungen
- 350 681 1 SMDokument5 Seiten350 681 1 SMBryan AbarcaNoch keine Bewertungen
- BiorremediacionDokument38 SeitenBiorremediacionGabriel Chambi AlanocaNoch keine Bewertungen
- Fitorremediación. Una Alternativa para Eliminar La Contaminación.Dokument19 SeitenFitorremediación. Una Alternativa para Eliminar La Contaminación.Ana paola Caballero fernandezNoch keine Bewertungen
- 12 - BiorremediaciónnDokument8 Seiten12 - BiorremediaciónnJuanGarciaBenavidesNoch keine Bewertungen
- La Biorremediacion en ColombiaDokument8 SeitenLa Biorremediacion en Colombiaoscar0590Noch keine Bewertungen
- Ejemplos de ToxicologiaDokument4 SeitenEjemplos de ToxicologiaIsrael José MendozaNoch keine Bewertungen
- Biorremediación Del SueloDokument4 SeitenBiorremediación Del SueloA CasNoch keine Bewertungen
- Biorremediacion Del Agua (F)Dokument15 SeitenBiorremediacion Del Agua (F)Josue LopezNoch keine Bewertungen
- Plaguicidas y contaminación ambientalDokument7 SeitenPlaguicidas y contaminación ambientalJavier Hernandez VelardeNoch keine Bewertungen
- Biorremediación de Suelos Contaminados Por Metales Pesados e HidrocarburosDokument9 SeitenBiorremediación de Suelos Contaminados Por Metales Pesados e HidrocarburosYanela ZeladaNoch keine Bewertungen
- 33173-Texto Del Artículo-197721-2-10-20140325Dokument17 Seiten33173-Texto Del Artículo-197721-2-10-20140325Fiore QuezadaNoch keine Bewertungen
- Ensayo La Ecologia y La BiotecnologiaDokument6 SeitenEnsayo La Ecologia y La BiotecnologiaStefany SilvaNoch keine Bewertungen
- Fitorremediacion en La Descontaminacion de Areas AgricolasDokument24 SeitenFitorremediacion en La Descontaminacion de Areas AgricolasRicsicitas BolivarNoch keine Bewertungen
- Fitorremediación: limpieza ambiental con plantasDokument8 SeitenFitorremediación: limpieza ambiental con plantasRoxana GuillènNoch keine Bewertungen
- Biorremediacion DiaposDokument70 SeitenBiorremediacion DiaposMaycol GutierrezNoch keine Bewertungen
- Biorremediacion de SuelosDokument17 SeitenBiorremediacion de SuelosNellyNoch keine Bewertungen
- BIORREMEDIACIÓNDokument22 SeitenBIORREMEDIACIÓNbetzayNoch keine Bewertungen
- Articulo - Biodegradacion Aerobia de Pesticidas HalogenadosDokument5 SeitenArticulo - Biodegradacion Aerobia de Pesticidas HalogenadosAlexa SolisNoch keine Bewertungen
- ARTICULO BASE 2 Biorremediacion Suelos Contaminados DDTDokument17 SeitenARTICULO BASE 2 Biorremediacion Suelos Contaminados DDTAngieNoch keine Bewertungen
- Biorremediacion-Microorganismos Que Limpian El AmbienteDokument10 SeitenBiorremediacion-Microorganismos Que Limpian El AmbientesofiNoch keine Bewertungen
- Tarea 3 Solucionalr La Contaminacion de PlaguicidaDokument4 SeitenTarea 3 Solucionalr La Contaminacion de PlaguicidaOscar julian MuñozNoch keine Bewertungen
- FitorecuperacionDokument16 SeitenFitorecuperacionWerner WernerNoch keine Bewertungen
- Remoción contaminantes colillas cigarro Pleurotus ostreatusDokument19 SeitenRemoción contaminantes colillas cigarro Pleurotus ostreatusAlan RamirezNoch keine Bewertungen
- Biorremediacion de Suelos Contaminados Por PetroleoDokument28 SeitenBiorremediacion de Suelos Contaminados Por PetroleoDayanaNoch keine Bewertungen
- Los Hongos Filamentosos, Su Importancia de Estudio y de Aplicación IndustrialDokument7 SeitenLos Hongos Filamentosos, Su Importancia de Estudio y de Aplicación IndustrialAlvaro CalderonNoch keine Bewertungen
- La Biorremediación Puede Ser Útil para La Degradación de Algunos ContaminantesDokument6 SeitenLa Biorremediación Puede Ser Útil para La Degradación de Algunos ContaminantesSofía DelgadoNoch keine Bewertungen
- IQM Fitorremediacion Biotec 25656 PDFDokument7 SeitenIQM Fitorremediacion Biotec 25656 PDFdeysiNoch keine Bewertungen
- Tarea 4 Proponer Solución Del Problema de Contaminación Del Suelo Escenario 2 - Version 2Dokument11 SeitenTarea 4 Proponer Solución Del Problema de Contaminación Del Suelo Escenario 2 - Version 2Yudis Ramirez OrozcoNoch keine Bewertungen
- El Cuaderno 36Dokument14 SeitenEl Cuaderno 36Katherine Cid RamírezNoch keine Bewertungen
- Elaborado Por:: Andres Carlosama Camilo Meneses Karina Pantoja Eider PortilloDokument27 SeitenElaborado Por:: Andres Carlosama Camilo Meneses Karina Pantoja Eider PortilloandresNoch keine Bewertungen
- BiorremediaciónDokument4 SeitenBiorremediaciónJ. Bravo RodriguezNoch keine Bewertungen
- El Cuaderno 36 - 1Dokument15 SeitenEl Cuaderno 36 - 1kiefer_hardNoch keine Bewertungen
- El Papel de Los Microorganismos en La Biodegradación de Compuestos TóxicosDokument6 SeitenEl Papel de Los Microorganismos en La Biodegradación de Compuestos TóxicosAlfredo CruzNoch keine Bewertungen
- TareaSem#7 DanielCastellanos 62111627Dokument8 SeitenTareaSem#7 DanielCastellanos 62111627Tops-5 GamesNoch keine Bewertungen
- XenobioticosDokument11 SeitenXenobioticosSaid Leonardo Cortez SandovalNoch keine Bewertungen
- Depuración Aguas Con PesticidasDokument10 SeitenDepuración Aguas Con PesticidasJesús Gómez DíazNoch keine Bewertungen
- AporteTarea4 ChristianBernalDokument7 SeitenAporteTarea4 ChristianBernalIbeth MuñozNoch keine Bewertungen
- BioremediaciónDokument4 SeitenBioremediaciónValentina HernándezNoch keine Bewertungen
- Tecnicas Biorremediacion - COMPLETODokument59 SeitenTecnicas Biorremediacion - COMPLETOMicaela MoránNoch keine Bewertungen
- Biorremediacion MicrobianaDokument4 SeitenBiorremediacion Microbianagonzalo pinNoch keine Bewertungen
- BIORREMEDIACIÓNDokument3 SeitenBIORREMEDIACIÓNMirtaNoch keine Bewertungen
- Contaminación del suelo por pesticidas y residuos industrialesDokument9 SeitenContaminación del suelo por pesticidas y residuos industrialesMaria Jose ChavarroNoch keine Bewertungen
- BIORREMEDIACIONDokument7 SeitenBIORREMEDIACIONEduardo Rosales CastroNoch keine Bewertungen
- Trabajo Final Completo - SueloDokument15 SeitenTrabajo Final Completo - SueloFRYDA AHIDALY LUJAN MENDEZNoch keine Bewertungen
- DDTDokument17 SeitenDDTDann Lup TooraLiin100% (1)
- Biosurfactantes y Su Papel en La Biorremediación de Suelos Contaminados Con PlaguicidasDokument21 SeitenBiosurfactantes y Su Papel en La Biorremediación de Suelos Contaminados Con PlaguicidasRicardo Ortiz GóngoraNoch keine Bewertungen
- FASE3 Propiedades y Contaminación Del SueloDokument16 SeitenFASE3 Propiedades y Contaminación Del SueloelmoralessNoch keine Bewertungen
- Biodegradación de compuestos xenobióticos y recalcitrantesDokument13 SeitenBiodegradación de compuestos xenobióticos y recalcitrantesSugeidys MoralesNoch keine Bewertungen
- AguasDokument33 SeitenAguasElizabeth KohatsuNoch keine Bewertungen
- LangostineraDokument13 SeitenLangostineraElizabeth KohatsuNoch keine Bewertungen
- Https de EconomiaDokument2 SeitenHttps de EconomiaElizabeth KohatsuNoch keine Bewertungen
- Vocabulario de MineraDokument8 SeitenVocabulario de MineraElizabeth KohatsuNoch keine Bewertungen
- Proyecto Terminado - 2 PDFDokument44 SeitenProyecto Terminado - 2 PDFElizabeth KohatsuNoch keine Bewertungen
- Maximas AvenidasDokument10 SeitenMaximas AvenidasElizabeth KohatsuNoch keine Bewertungen
- Flujo ModeloDokument1 SeiteFlujo ModeloElizabeth KohatsuNoch keine Bewertungen
- Matriz SeminarioDokument4 SeitenMatriz SeminarioElizabeth KohatsuNoch keine Bewertungen
- Proced I Mien ToDokument1 SeiteProced I Mien ToElizabeth KohatsuNoch keine Bewertungen
- Ordenacion ContingenteDokument14 SeitenOrdenacion ContingenteElizabeth KohatsuNoch keine Bewertungen
- Eglamour SlidesCarnivalDokument60 SeitenEglamour SlidesCarnivalElizabeth KohatsuNoch keine Bewertungen
- Precios UnitariosDokument12 SeitenPrecios UnitariosElizabeth KohatsuNoch keine Bewertungen
- Qui Ruvi LcaDokument1 SeiteQui Ruvi LcaElizabeth KohatsuNoch keine Bewertungen
- Obj - Int-ConDokument2 SeitenObj - Int-ConElizabeth KohatsuNoch keine Bewertungen
- Peso Inicial RudaDokument1 SeitePeso Inicial RudaElizabeth KohatsuNoch keine Bewertungen
- RELLENOS SANI y BOTADEROSDokument3 SeitenRELLENOS SANI y BOTADEROSIván De La Cruz CallupeNoch keine Bewertungen
- Sesion 1 ArcgisDokument3 SeitenSesion 1 ArcgisElizabeth KohatsuNoch keine Bewertungen
- Ataque QuimicoDokument2 SeitenAtaque QuimicoElizabeth KohatsuNoch keine Bewertungen
- VitisDokument24 SeitenVitisElizabeth KohatsuNoch keine Bewertungen
- Arcgis-Int-Sesion 1-Tarea-1.1Dokument2 SeitenArcgis-Int-Sesion 1-Tarea-1.1Elizabeth KohatsuNoch keine Bewertungen
- Identificacion de Carbohidratos A TravesDokument16 SeitenIdentificacion de Carbohidratos A TravesElizabeth KohatsuNoch keine Bewertungen
- Sesion 1 ArcgisDokument3 SeitenSesion 1 ArcgisElizabeth KohatsuNoch keine Bewertungen
- Arcgis-Bas-Sesion 10-Tarea-1.1Dokument3 SeitenArcgis-Bas-Sesion 10-Tarea-1.1Elizabeth Kohatsu0% (1)
- Examen de SIG-TB - BDokument1 SeiteExamen de SIG-TB - BElizabeth KohatsuNoch keine Bewertungen
- NestleDokument3 SeitenNestleElizabeth KohatsuNoch keine Bewertungen
- Atmos F EricaDokument4 SeitenAtmos F EricaElizabeth KohatsuNoch keine Bewertungen
- Tarea Arcgis Bas s2 Galvez KohatsuDokument5 SeitenTarea Arcgis Bas s2 Galvez KohatsuElizabeth Kohatsu50% (4)
- Linda Esta 2Dokument8 SeitenLinda Esta 2Elizabeth KohatsuNoch keine Bewertungen
- HUSKY PowerWasher 1750psi Operator's ManualDokument32 SeitenHUSKY PowerWasher 1750psi Operator's ManualCarter AtkinsonNoch keine Bewertungen
- Automatizacion y Control ProyectoDokument150 SeitenAutomatizacion y Control ProyectoMonse De MaslowNoch keine Bewertungen
- Cat Compre Sore SirDokument45 SeitenCat Compre Sore SirGiovanni RiveraNoch keine Bewertungen
- Análisis de Objeto Técnico La EscaleraDokument8 SeitenAnálisis de Objeto Técnico La EscaleraIng. Gerardo Sánchez Nájera100% (2)
- Informe Nº6 Turbinas PeltonDokument11 SeitenInforme Nº6 Turbinas PeltonRenzo Arone SolarNoch keine Bewertungen
- VFD FLEXIBLE VF - Ne-Coperint-EsDokument2 SeitenVFD FLEXIBLE VF - Ne-Coperint-Esjorgema34Noch keine Bewertungen
- Insert Lea Ps KW 10863993 AssemblysheetDokument2 SeitenInsert Lea Ps KW 10863993 AssemblysheetSergio Matador VegaNoch keine Bewertungen
- PresupuestoConstrucciónViviendaDokument18 SeitenPresupuestoConstrucciónViviendaAldair PJNoch keine Bewertungen
- Banqueo ECU Toyota 4EDokument7 SeitenBanqueo ECU Toyota 4ELuis TarquiNoch keine Bewertungen
- Método de Hunsaker y RightmireDokument3 SeitenMétodo de Hunsaker y RightmireAlvaro Eduardo Barrantes ZelaNoch keine Bewertungen
- Recomendaciones Santa Cruz de La SierraDokument15 SeitenRecomendaciones Santa Cruz de La SierrajpjazzpianoNoch keine Bewertungen
- Informe de SedimentaciónDokument15 SeitenInforme de SedimentaciónCarolina Arias Gallego0% (1)
- Modelamiento de Gases LadrilleraDokument19 SeitenModelamiento de Gases LadrilleraKatherine ZuñigaNoch keine Bewertungen
- RESUMENDokument69 SeitenRESUMENKathya milena Sandoval AlcalaNoch keine Bewertungen
- RICOH Pro5100s PDFDokument8 SeitenRICOH Pro5100s PDFquiquescribdNoch keine Bewertungen
- Lab NORMAS DE SEGURIDADDokument12 SeitenLab NORMAS DE SEGURIDADorlandoNoch keine Bewertungen
- Marco Teórico FoproiDokument16 SeitenMarco Teórico FoproiJoan CamposNoch keine Bewertungen
- Señales de tránsitoDokument7 SeitenSeñales de tránsitoMerlin Asprilla AnguloNoch keine Bewertungen
- Ejercicio 1 de HerramientasDokument2 SeitenEjercicio 1 de HerramientasWilmer Castope RuizNoch keine Bewertungen
- Cuestionario de Las Principales Aleaciones No FerrosasDokument4 SeitenCuestionario de Las Principales Aleaciones No FerrosasHectorNoch keine Bewertungen
- Cuestionario 3 TopografiaDokument4 SeitenCuestionario 3 TopografiaPuro EntretenimientoNoch keine Bewertungen
- Tarea 3Dokument8 SeitenTarea 3Jesus Fabian ArenasNoch keine Bewertungen