Das könnte Ihnen auch gefallen
- Design Constraints of Horizontal SeparatorDokument2 SeitenDesign Constraints of Horizontal SeparatorBadzlinaKhairunizzahraNoch keine Bewertungen
- Bab 3 - Ranya - RevisiDokument4 SeitenBab 3 - Ranya - RevisiBadzlinaKhairunizzahraNoch keine Bewertungen
- Design Contraints BadzDokument2 SeitenDesign Contraints BadzBadzlinaKhairunizzahraNoch keine Bewertungen
- Scan Data PreliumDokument5 SeitenScan Data PreliumBadzlinaKhairunizzahraNoch keine Bewertungen
- Anti Stain Deodorant: Group 25Dokument13 SeitenAnti Stain Deodorant: Group 25BadzlinaKhairunizzahraNoch keine Bewertungen
- Table X. Communication Cost Component Cost/month Cost/yearDokument15 SeitenTable X. Communication Cost Component Cost/month Cost/yearBadzlinaKhairunizzahraNoch keine Bewertungen
- Chemical Engineering Department, Faculty of Engineering, Universitas Indonesia, Depok, West Java, IndonesiaDokument6 SeitenChemical Engineering Department, Faculty of Engineering, Universitas Indonesia, Depok, West Java, IndonesiaBadzlinaKhairunizzahraNoch keine Bewertungen
- POLYMATH Results: Multiple Linear RegressionDokument1 SeitePOLYMATH Results: Multiple Linear RegressionBadzlinaKhairunizzahraNoch keine Bewertungen
- Surface Facilities 2 PDFDokument54 SeitenSurface Facilities 2 PDFBadzlinaKhairunizzahraNoch keine Bewertungen
- UopDokument8 SeitenUopBadzlinaKhairunizzahraNoch keine Bewertungen
- Final Report - BadzDokument36 SeitenFinal Report - BadzBadzlinaKhairunizzahraNoch keine Bewertungen
- Concept ScoringDokument4 SeitenConcept ScoringBadzlinaKhairunizzahraNoch keine Bewertungen
- POLYMATH Results: Multiple Linear RegressionDokument1 SeitePOLYMATH Results: Multiple Linear RegressionBadzlinaKhairunizzahraNoch keine Bewertungen
- Cipro Flax inDokument4 SeitenCipro Flax inBadzlinaKhairunizzahraNoch keine Bewertungen
- LuncheonDokument1 SeiteLuncheonBadzlinaKhairunizzahraNoch keine Bewertungen
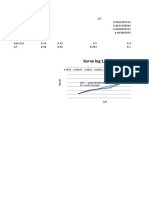
- Kurva Log 1/t Vs 1/TDokument3 SeitenKurva Log 1/t Vs 1/TBadzlinaKhairunizzahraNoch keine Bewertungen
- Analisis Farmasi Dasar - 1 REVDokument126 SeitenAnalisis Farmasi Dasar - 1 REVBadzlinaKhairunizzahra100% (1)
- Uas Practice I. Structure (25 Points) A. Use ACTIVE or PASSIVE, in Any Appropriate Tense, For The Verbs in Parentheses (15 Points)Dokument7 SeitenUas Practice I. Structure (25 Points) A. Use ACTIVE or PASSIVE, in Any Appropriate Tense, For The Verbs in Parentheses (15 Points)BadzlinaKhairunizzahraNoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Pedia Allergic DisordersDokument4 SeitenPedia Allergic Disordersmia liaNoch keine Bewertungen
- Д аминдэм дархлаа тогтолцоонд оролцох нь PDFDokument10 SeitenД аминдэм дархлаа тогтолцоонд оролцох нь PDFЭ.ТөгөлдөрNoch keine Bewertungen
- Pathology Mcqs CorrectDokument45 SeitenPathology Mcqs CorrectAhmed Alamin83% (6)
- Mehu525 U4 T3 INTOXICACIONES2-4Dokument32 SeitenMehu525 U4 T3 INTOXICACIONES2-4jorgealonsosaavedraNoch keine Bewertungen
- Difference Between Mast Cell and BasophilDokument2 SeitenDifference Between Mast Cell and BasophilponbohacopNoch keine Bewertungen
- Greyson 2019Dokument10 SeitenGreyson 2019Ziha Zia Leonita FauziNoch keine Bewertungen
- Hypereosinophilic Sydrome in 2 CatsDokument5 SeitenHypereosinophilic Sydrome in 2 Catscnicipor4873Noch keine Bewertungen
- Parasitology Study GuideDokument16 SeitenParasitology Study GuideesraaNoch keine Bewertungen
- Novel Anti-Inflammatory Treatments For Asthma: ReviewDokument13 SeitenNovel Anti-Inflammatory Treatments For Asthma: Reviewnopera kurnia watiNoch keine Bewertungen
- Wa0005.Dokument4 SeitenWa0005.science worldNoch keine Bewertungen
- Hypersensitivity AssignmentDokument6 SeitenHypersensitivity AssignmentNicholasNoch keine Bewertungen
- Chapter 15 Lung Pathology ErnieDokument20 SeitenChapter 15 Lung Pathology ErnieZandra Lyn AlundayNoch keine Bewertungen
- Helmintos NematodosDokument12 SeitenHelmintos NematodosHANNIANoch keine Bewertungen
- UW Allergy + Immuno - Educational Objectives PDFDokument25 SeitenUW Allergy + Immuno - Educational Objectives PDFDrbee10Noch keine Bewertungen
- Endo-Perio Lesion PDFDokument23 SeitenEndo-Perio Lesion PDFamaniNoch keine Bewertungen
- Immunology For Pharmacy Students PDFDokument199 SeitenImmunology For Pharmacy Students PDFandirio7486Noch keine Bewertungen
- Hamza DissertDokument108 SeitenHamza DissertMobin Ur Rehman KhanNoch keine Bewertungen
- Really Essential Medical ImmunologyDokument224 SeitenReally Essential Medical Immunology99noname100% (2)
- Management of Allergic Rhinitis: P. Van Cauwenberge, H. Van HoeckeDokument20 SeitenManagement of Allergic Rhinitis: P. Van Cauwenberge, H. Van HoeckePeter SalimNoch keine Bewertungen
- Atlas of Clinical HematologyDokument146 SeitenAtlas of Clinical HematologyMaath Khalid100% (11)
- 2002, Vol.86, Issues 5, AsthmaDokument229 Seiten2002, Vol.86, Issues 5, AsthmaFaisal H RanaNoch keine Bewertungen
- GRANULOPEISISDokument39 SeitenGRANULOPEISISkemalmiaNoch keine Bewertungen
- LECTURE 1 HANDOUT Online ImmuneDokument28 SeitenLECTURE 1 HANDOUT Online ImmuneFYMNoch keine Bewertungen
- Immunedermatology MCQsDokument73 SeitenImmunedermatology MCQsDr.Tawheed90% (10)
- Heamatology 1Dokument57 SeitenHeamatology 1Victoria &BeautifulPeopleNoch keine Bewertungen
- Immunotoxicology 2007dll-2Dokument80 SeitenImmunotoxicology 2007dll-2Puyi PuyenkNoch keine Bewertungen
- Eosinophil Count - Explanation & RisksDokument9 SeitenEosinophil Count - Explanation & RisksvinodNoch keine Bewertungen
- Hema Lec Prefinals ExamDokument18 SeitenHema Lec Prefinals ExamMark jay LlanoNoch keine Bewertungen
- The Blood: Rubie Maranan-Causaren, MSDokument73 SeitenThe Blood: Rubie Maranan-Causaren, MSlady ann jimenez100% (3)
- Functional Gastrointestinal Disorders 3: SeriesDokument14 SeitenFunctional Gastrointestinal Disorders 3: SeriesDimas RizkyNoch keine Bewertungen