Das könnte Ihnen auch gefallen
- Zanphorlin 2011Dokument7 SeitenZanphorlin 2011Arturo PeñablassiNoch keine Bewertungen
- Papain-Like Peptidases: Structure, Function, and Evolution: Marko Novinec and Brigita Lenar Č I ČDokument22 SeitenPapain-Like Peptidases: Structure, Function, and Evolution: Marko Novinec and Brigita Lenar Č I Čpedro augustoNoch keine Bewertungen
- Adaptive Evolution and Functional Divergence of Pepsin Gene FamilyDokument10 SeitenAdaptive Evolution and Functional Divergence of Pepsin Gene FamilyharisankarhsNoch keine Bewertungen
- Pahala 1Dokument5 SeitenPahala 1Daeng MangkaNoch keine Bewertungen
- Cytotoxic Effects of Repin, A Principal Sesquiterpene Lactone of Russian KnapweedDokument8 SeitenCytotoxic Effects of Repin, A Principal Sesquiterpene Lactone of Russian KnapweedcthjgfytvfnoNoch keine Bewertungen
- 2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureDokument10 Seiten2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureIasmin ResendeNoch keine Bewertungen
- A New Group of Aromatic Prenyltransferases in Fungi, Catalyzing A 2,7-Dihydroxynaphthalene J. Biol. Chem.-2010-Haug-Schifferdecker-16487-94Dokument9 SeitenA New Group of Aromatic Prenyltransferases in Fungi, Catalyzing A 2,7-Dihydroxynaphthalene J. Biol. Chem.-2010-Haug-Schifferdecker-16487-94ckmarloweNoch keine Bewertungen
- Balasubramanian 2013 Cloning and Molecular Analysis of The Aspartic Protease Sc-ASP110 GeneDokument8 SeitenBalasubramanian 2013 Cloning and Molecular Analysis of The Aspartic Protease Sc-ASP110 GeneIliana Zazueta OjedaNoch keine Bewertungen
- Jsir 61 (9) 690-704 PDFDokument15 SeitenJsir 61 (9) 690-704 PDFAusteridad LopezNoch keine Bewertungen
- Kay, J. (N.D.) - Aspartic Proteinases and Their Inhibitors. Aspartic Proteinases and Their Inhibitors, 1-18.Dokument18 SeitenKay, J. (N.D.) - Aspartic Proteinases and Their Inhibitors. Aspartic Proteinases and Their Inhibitors, 1-18.MauNoch keine Bewertungen
- Use Only: New in Vitro Dna Polymerase Activity and Fidelity Assay Reveals Age-Dependent Changes in Arabidopsis ThalianaDokument6 SeitenUse Only: New in Vitro Dna Polymerase Activity and Fidelity Assay Reveals Age-Dependent Changes in Arabidopsis ThalianacroitorugeorgianaNoch keine Bewertungen
- EnzymesDokument13 SeitenEnzymesManjusha KondepudiNoch keine Bewertungen
- Naringenina 3 International Review of Cell and Molecular BiologyDokument13 SeitenNaringenina 3 International Review of Cell and Molecular BiologyCristofer Perez SeguraNoch keine Bewertungen
- Influence of antidepressants on salivary gland ecto-enzymesDokument7 SeitenInfluence of antidepressants on salivary gland ecto-enzymesconstanceNoch keine Bewertungen
- 1 FullDokument9 Seiten1 FullFauziana NurhanisahNoch keine Bewertungen
- TMP EAD3Dokument21 SeitenTMP EAD3FrontiersNoch keine Bewertungen
- Choi 2012Dokument6 SeitenChoi 2012Yudho AdhitomoNoch keine Bewertungen
- Bioorganic & Medicinal Chemistry LettersDokument5 SeitenBioorganic & Medicinal Chemistry LettersElinaNoch keine Bewertungen
- Jurnal Konsentrasi Pegagan 2.5, 5.0 and 10.0 MGMLDokument7 SeitenJurnal Konsentrasi Pegagan 2.5, 5.0 and 10.0 MGMLNurvia AndrianiNoch keine Bewertungen
- (18469558 - Acta Pharmaceutica) Anthelmintic Activity of Praziquantel and Spilanthes Acmella Extract On An Intestinal Cestode ParasiteDokument10 Seiten(18469558 - Acta Pharmaceutica) Anthelmintic Activity of Praziquantel and Spilanthes Acmella Extract On An Intestinal Cestode ParasiteDiego NamucheNoch keine Bewertungen
- DNA Repair Protein Involved in Heart and Blood DevelopmentDokument12 SeitenDNA Repair Protein Involved in Heart and Blood DevelopmentSol Jumaide WerbleNoch keine Bewertungen
- Biochemical analysis of a papain-like protease from Asclepias curassavica latexDokument9 SeitenBiochemical analysis of a papain-like protease from Asclepias curassavica latexPavan KNoch keine Bewertungen
- A Molecular Description of Acid PhosphataseDokument24 SeitenA Molecular Description of Acid Phosphatasekawtarmar3Noch keine Bewertungen
- Identification, Enzymatic Spoilage Characterization and ProteolyticDokument12 SeitenIdentification, Enzymatic Spoilage Characterization and ProteolyticCristhian JohnNoch keine Bewertungen
- Ijms 20 00107Dokument20 SeitenIjms 20 00107SRI RAHMAYUNINoch keine Bewertungen
- 2019 Article 421DTNDokument8 Seiten2019 Article 421DTNDulce Maria AngelesNoch keine Bewertungen
- 2008 BBA-Lingaraju and GowdaDokument7 Seiten2008 BBA-Lingaraju and GowdaLalitha R GowdaNoch keine Bewertungen
- 02 Insect Biochemistry MolecularDokument0 Seiten02 Insect Biochemistry Molecularlalo199Noch keine Bewertungen
- MainDokument7 SeitenMainLeena John MathaiNoch keine Bewertungen
- 2011 Article 9671Dokument9 Seiten2011 Article 9671Brandon GriffinNoch keine Bewertungen
- Protease: Hierarchy of ProteasesDokument9 SeitenProtease: Hierarchy of ProteasesCrow LordNoch keine Bewertungen
- Chen 2016Dokument9 SeitenChen 2016Shampa SenNoch keine Bewertungen
- Enzymatic Determination of Catechol Oxidase and Protease From Fruits (Orange, Apple) and Vegetables (Carrot, Tomato)Dokument7 SeitenEnzymatic Determination of Catechol Oxidase and Protease From Fruits (Orange, Apple) and Vegetables (Carrot, Tomato)International Organization of Scientific Research (IOSR)Noch keine Bewertungen
- Serum biomarkers predict risk of retained placenta in dairy cowsDokument2 SeitenSerum biomarkers predict risk of retained placenta in dairy cowsHakim HallouzNoch keine Bewertungen
- Tobacco Etch Virus Protease: A Shortcut Across BiotechnologiesDokument16 SeitenTobacco Etch Virus Protease: A Shortcut Across BiotechnologiesNhật ThiệnNoch keine Bewertungen
- Protective Effect of Casperome An Orally Bioavailable Frankincense Extract On Lipopolysaccharide Induced Systemic Inflammation in MiceDokument18 SeitenProtective Effect of Casperome An Orally Bioavailable Frankincense Extract On Lipopolysaccharide Induced Systemic Inflammation in Miceana mariaNoch keine Bewertungen
- A Comparative Study On Cell Disruption Methods PDFDokument8 SeitenA Comparative Study On Cell Disruption Methods PDFMarisol MancillaNoch keine Bewertungen
- Camara Et Al 2017 (Mattanovich GRP) Increased Dosage of AOX1promoter Transcription AttenuationDokument16 SeitenCamara Et Al 2017 (Mattanovich GRP) Increased Dosage of AOX1promoter Transcription AttenuationAnaNoch keine Bewertungen
- Fosfatase Soluvel e LeismaniaDokument8 SeitenFosfatase Soluvel e LeismaniacoringacravoNoch keine Bewertungen
- Differential Expression Profiling of The Hepatic Proteome in A Rat Model of Dioxin ResistanceDokument13 SeitenDifferential Expression Profiling of The Hepatic Proteome in A Rat Model of Dioxin Resistancegiasco78Noch keine Bewertungen
- Shu - 2009 Aspergillus EstruturaDokument8 SeitenShu - 2009 Aspergillus EstruturaTatiane RibeiroNoch keine Bewertungen
- Shi 2015Dokument11 SeitenShi 2015Bruno FreireNoch keine Bewertungen
- TMP C039Dokument5 SeitenTMP C039FrontiersNoch keine Bewertungen
- Protease Inhibitors: Assignment OnDokument6 SeitenProtease Inhibitors: Assignment OnVinod Kumar0% (1)
- Caracteres y Proteinas en A. PisumDokument16 SeitenCaracteres y Proteinas en A. PisumJhonny Andre Blas PonceNoch keine Bewertungen
- Extracellular Expression of Alkaline Phytase in Pichia Pastoris InfluenceDokument7 SeitenExtracellular Expression of Alkaline Phytase in Pichia Pastoris InfluenceNatalia Andrea Gómez RaveNoch keine Bewertungen
- In Vitro Effects of Thai Medicinal Plants On Human Lymphocyte Activity PDFDokument8 SeitenIn Vitro Effects of Thai Medicinal Plants On Human Lymphocyte Activity PDFmadelineNoch keine Bewertungen
- Identification and Characterisation of Gut Proteases in The Fig Tree Skeletoniser Moth, Choreutis NemoranaDokument8 SeitenIdentification and Characterisation of Gut Proteases in The Fig Tree Skeletoniser Moth, Choreutis NemoranaRositaNoch keine Bewertungen
- TMP 5 EC8Dokument12 SeitenTMP 5 EC8FrontiersNoch keine Bewertungen
- البحث السادسDokument11 Seitenالبحث السادسsoha ahmedNoch keine Bewertungen
- Review of Pentatricopeptide Repeat Proteins: A Socket Set for Organelle Gene ExpressionDokument8 SeitenReview of Pentatricopeptide Repeat Proteins: A Socket Set for Organelle Gene ExpressionPavithra MohanNoch keine Bewertungen
- 1 s2.0 S2211505612001044 MainDokument5 Seiten1 s2.0 S2211505612001044 MainMichaelNoch keine Bewertungen
- Many of The Slides Are Downloaded From Google ImagesDokument51 SeitenMany of The Slides Are Downloaded From Google ImagesAysha MohideenNoch keine Bewertungen
- Purine and Pyrimidine Nucleotide Synthesis and MetDokument21 SeitenPurine and Pyrimidine Nucleotide Synthesis and MetAbdul Matin RahimiNoch keine Bewertungen
- Abranches 2009Dokument9 SeitenAbranches 2009Emma BouveretNoch keine Bewertungen
- Journal of Plant Physiology: Yee-Min Kwan, Sariah Meon, Chai-Ling Ho, Mui-Yun WongDokument6 SeitenJournal of Plant Physiology: Yee-Min Kwan, Sariah Meon, Chai-Ling Ho, Mui-Yun WongroseNoch keine Bewertungen
- Neurotrophic Properties of The Lion's Mane Medicinal Mushroom, Hericium Erinaceus (Higher Basidiomycetes) From MalaysiaDokument16 SeitenNeurotrophic Properties of The Lion's Mane Medicinal Mushroom, Hericium Erinaceus (Higher Basidiomycetes) From MalaysiaDayon BochniaNoch keine Bewertungen
- NIH Public Access: Author ManuscriptDokument21 SeitenNIH Public Access: Author ManuscriptAlberto Aguilar BriseñoNoch keine Bewertungen
- Serine Protease and Serine Protease Inhibitors: Advanced ArticleDokument8 SeitenSerine Protease and Serine Protease Inhibitors: Advanced ArticleqhqhqNoch keine Bewertungen
- Food Research InternationalDokument10 SeitenFood Research InternationallandangeNoch keine Bewertungen
- 2016-Proteomic Analysis and Purification of An Unusual Germin-LikeDokument14 Seiten2016-Proteomic Analysis and Purification of An Unusual Germin-LikelandangeNoch keine Bewertungen
- Dairy Science and Technology (CRC 2005)Dokument8 SeitenDairy Science and Technology (CRC 2005)landangeNoch keine Bewertungen
- FirstChoice RLM-RACE Kit Manual AnnotatedDokument33 SeitenFirstChoice RLM-RACE Kit Manual AnnotatedlandangeNoch keine Bewertungen
- Inclusion Body Solubilization ReagentDokument2 SeitenInclusion Body Solubilization ReagentlandangeNoch keine Bewertungen
- 1 s2.0 S0278691515001179 MainDokument11 Seiten1 s2.0 S0278691515001179 MainlandangeNoch keine Bewertungen
- Refolding Out of Guanidine Hydrochloride Is AnDokument8 SeitenRefolding Out of Guanidine Hydrochloride Is AnlandangeNoch keine Bewertungen
- A Novel Protein Refolding Protocol For The Solubilization and PurificationDokument6 SeitenA Novel Protein Refolding Protocol For The Solubilization and PurificationlandangeNoch keine Bewertungen
- Mechanism and Model For Solubilization of Inclusion BodiesDokument11 SeitenMechanism and Model For Solubilization of Inclusion BodieslandangeNoch keine Bewertungen
- CD Review-2009 DicroismoDokument20 SeitenCD Review-2009 DicroismolandangeNoch keine Bewertungen
- The Effects of Glycerol, Glucose, GaDokument8 SeitenThe Effects of Glycerol, Glucose, GalandangeNoch keine Bewertungen
- Nivel SIII 2008.2 PDFDokument3 SeitenNivel SIII 2008.2 PDFlandangeNoch keine Bewertungen
- A Cysteine Protease Isolated From The Latex of FicusDokument13 SeitenA Cysteine Protease Isolated From The Latex of FicuslandangeNoch keine Bewertungen
- EP0588326A2Dokument9 SeitenEP0588326A2Mohsen GolmohammadiNoch keine Bewertungen
- Molecular Basis of InheritanceDokument37 SeitenMolecular Basis of Inheritancechristopher lopezNoch keine Bewertungen
- EFFECTS OF RADIATION ON PLANTSDokument35 SeitenEFFECTS OF RADIATION ON PLANTSOkaroFrankNoch keine Bewertungen
- Sugar Mills Presention, by Aziz Jamali TalharDokument25 SeitenSugar Mills Presention, by Aziz Jamali TalharAnonymous v32yhQWVFNoch keine Bewertungen
- Smartlab - Bahan Kimia PDFDokument2 SeitenSmartlab - Bahan Kimia PDFPandeglang laboratoriumNoch keine Bewertungen
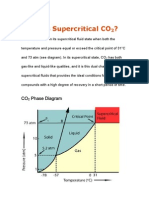
- What Is Super Critical CO2Dokument42 SeitenWhat Is Super Critical CO2kesaco100% (2)
- D2000-2018 Standard Classification System For Rubber Products in Automotive ApplicationsDokument53 SeitenD2000-2018 Standard Classification System For Rubber Products in Automotive ApplicationsLuisDupuy67% (3)
- Cy1002 PDFDokument36 SeitenCy1002 PDFAryan PandeyNoch keine Bewertungen
- Synthesis of Grafted Polylactic Acid and Polyhydroxyalkanoate by A Green Reactive Extrusion ProcessDokument15 SeitenSynthesis of Grafted Polylactic Acid and Polyhydroxyalkanoate by A Green Reactive Extrusion ProcessIrma BrennanNoch keine Bewertungen
- Aminoethyl Ethanolamine - AEEA: Product InformationDokument1 SeiteAminoethyl Ethanolamine - AEEA: Product InformationtotpityiNoch keine Bewertungen
- INCOMPATIBILITY CAUSESDokument40 SeitenINCOMPATIBILITY CAUSESnizam_ghaniNoch keine Bewertungen
- Astm D0330 - 1 (En)Dokument2 SeitenAstm D0330 - 1 (En)svvasin2013Noch keine Bewertungen
- Process Economics Program - Pta ProcessDokument10 SeitenProcess Economics Program - Pta ProcessvasucristalNoch keine Bewertungen
- Fatty Acid MetabolismDokument28 SeitenFatty Acid MetabolismMitchelle SaurambaNoch keine Bewertungen
- 9701 s07 Ms 4Dokument8 Seiten9701 s07 Ms 4Vivian SiewNoch keine Bewertungen
- Introduction To Composite Materials SlidesDokument343 SeitenIntroduction To Composite Materials SlideseduserenzaNoch keine Bewertungen
- CARBOHYDRATESDokument38 SeitenCARBOHYDRATESgulrukh100% (3)
- Eviar 100% Waterproof Flooring CatalogueDokument11 SeitenEviar 100% Waterproof Flooring CatalogueKelly YuenNoch keine Bewertungen
- SNS Grandes ComerciosDokument24 SeitenSNS Grandes ComerciosMaximiliano ToscanoNoch keine Bewertungen
- 2020 ACJC H2 Paper 3 AnswersDokument23 Seiten2020 ACJC H2 Paper 3 AnswersbeverlyyyNoch keine Bewertungen
- The Biological Importance of Water: Essential for All LifeDokument1 SeiteThe Biological Importance of Water: Essential for All LifesayraNoch keine Bewertungen
- Biobutanol - Advantages and Disavantages. Comparision To Ethanol.Dokument9 SeitenBiobutanol - Advantages and Disavantages. Comparision To Ethanol.Phi TiêuNoch keine Bewertungen
- INDUSTRIAL INTERNSHIP REPORT ON MANUFACTURING OF DYESDokument50 SeitenINDUSTRIAL INTERNSHIP REPORT ON MANUFACTURING OF DYES1031 Ayush NasitNoch keine Bewertungen
- IITJEE | MEDICAL | Question Bank On S-Block ElementsDokument7 SeitenIITJEE | MEDICAL | Question Bank On S-Block ElementsAshutosh TripathiNoch keine Bewertungen
- (WWW - Entrance-Exam - Net) - ICAR ARS NET Previous Questions On Agricultural Biotechnology 1Dokument5 Seiten(WWW - Entrance-Exam - Net) - ICAR ARS NET Previous Questions On Agricultural Biotechnology 1ArvindUpadhyayNoch keine Bewertungen
- 7.1 - Nucleic Acids, Ahl: (Adapted From)Dokument5 Seiten7.1 - Nucleic Acids, Ahl: (Adapted From)Big CinemaNoch keine Bewertungen
- Astm C582 (1995) PDFDokument7 SeitenAstm C582 (1995) PDFamusumuseeNoch keine Bewertungen
- Method Statement - Terrace WP - 08.07Dokument4 SeitenMethod Statement - Terrace WP - 08.07Treon Ajmera100% (2)
- Quick Revision-Chem - Ssce 2023Dokument3 SeitenQuick Revision-Chem - Ssce 2023Jampa SaicharanNoch keine Bewertungen