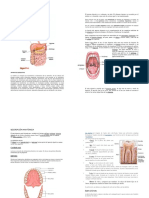
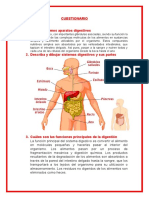
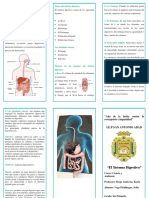
Das könnte Ihnen auch gefallen
- Lección 3 Supervisor de OperacionesDokument16 SeitenLección 3 Supervisor de OperacionesBrayhan Sánchez67% (6)
- Nivel 3 Lección 1Dokument5 SeitenNivel 3 Lección 1Brayhan Sánchez0% (2)
- Encuesta Miel de AbejaDokument5 SeitenEncuesta Miel de AbejaGradiz Murillo100% (2)
- DIAGRAMA DEL FLUJO DE PROCESO DE DESMONTAJEDokument2 SeitenDIAGRAMA DEL FLUJO DE PROCESO DE DESMONTAJEBrayhan Sánchez100% (1)
- 6toinforme Movilidad Como Vamos Lima PDFDokument12 Seiten6toinforme Movilidad Como Vamos Lima PDFJorge Luis Lumbre Santa CruzNoch keine Bewertungen
- Frenos Jacobs 404 en l10 Cummins PDFDokument23 SeitenFrenos Jacobs 404 en l10 Cummins PDFJuan Carlos SoHeNoch keine Bewertungen
- Análisis microscópico intestino rata H/EDokument8 SeitenAnálisis microscópico intestino rata H/EMilagros JofreNoch keine Bewertungen
- Comparación Morfofisiológica de Tractos Digestivos de Animales y Sus Estructuras AlimentariasDokument13 SeitenComparación Morfofisiológica de Tractos Digestivos de Animales y Sus Estructuras AlimentariasMaria Del Carmen Pimienta SolanoNoch keine Bewertungen
- Guía Nutrición en AnimalesDokument4 SeitenGuía Nutrición en AnimalesAdrianaPolancoGonzálezNoch keine Bewertungen
- DigestivoDokument6 SeitenDigestivomauricio jose Loreto aponteNoch keine Bewertungen
- Gene Aparato DigestivoDokument1 SeiteGene Aparato DigestivoPablo Vega MNoch keine Bewertungen
- Sistema digestivo rumiantesDokument5 SeitenSistema digestivo rumiantesMarco Sebastian Benites CruzNoch keine Bewertungen
- Aparato DigestivoDokument8 SeitenAparato DigestivoyorgelisNoch keine Bewertungen
- Sistema Digestivo y RenalDokument5 SeitenSistema Digestivo y RenalAleNoch keine Bewertungen
- Guia de Sistema DigestivoDokument9 SeitenGuia de Sistema DigestivoCarlos Irigoin JaraNoch keine Bewertungen
- Fisiología AviarDokument29 SeitenFisiología AviarLuis David VillanuevaNoch keine Bewertungen
- Resumen Sist - GastrointestinalDokument21 SeitenResumen Sist - GastrointestinalMaria Jose FloresNoch keine Bewertungen
- 4 Unidad de Fisiologia AnimalDokument11 Seiten4 Unidad de Fisiologia AnimalJuana LópezNoch keine Bewertungen
- Nutricion Animal Módulo 2Dokument10 SeitenNutricion Animal Módulo 2IariitaGordilloNoch keine Bewertungen
- El Aparato DigestivoDokument7 SeitenEl Aparato DigestivoJorge BravoNoch keine Bewertungen
- Sistema Del Cuerpo HumanoDokument7 SeitenSistema Del Cuerpo HumanotammyNoch keine Bewertungen
- Fisiología digestiva en perros y gatosDokument1 SeiteFisiología digestiva en perros y gatosJessNoch keine Bewertungen
- El Sistema DigestivoDokument4 SeitenEl Sistema DigestivoJuan NavasNoch keine Bewertungen
- Digestión-Animal-para-Tercer-y 4toGrado-de-Secundaria InvertebradosDokument4 SeitenDigestión-Animal-para-Tercer-y 4toGrado-de-Secundaria Invertebradosgustavo gonzaleschavezNoch keine Bewertungen
- TEMA 13 Gulag FreeDokument29 SeitenTEMA 13 Gulag FreeLuciaNoch keine Bewertungen
- Tema 4Dokument4 SeitenTema 4Sara Gamboa JuradoNoch keine Bewertungen
- Taller Funcion de NutricionDokument6 SeitenTaller Funcion de Nutricionlaura50% (2)
- CsNat - Cap11.s en Línea.Dokument16 SeitenCsNat - Cap11.s en Línea.Carola JamesNoch keine Bewertungen
- Tarea 2Dokument6 SeitenTarea 2Emelyn Paola Calderon DuranNoch keine Bewertungen
- BIO4 CAP2 ActivadosDokument14 SeitenBIO4 CAP2 ActivadosCarola JamesNoch keine Bewertungen
- Tema 7 Anatomia Del Aparato DigestivoDokument15 SeitenTema 7 Anatomia Del Aparato DigestivoYodyNoch keine Bewertungen
- 7-3 Biologia Guia 05 Per 1Dokument6 Seiten7-3 Biologia Guia 05 Per 1Bibiana Montero InsuastyNoch keine Bewertungen
- Sistema Digestivo HarrisonDokument228 SeitenSistema Digestivo HarrisonJ. JaimesNoch keine Bewertungen
- Repaso Evaluación Santillana Ciencias Naturales 5Dokument12 SeitenRepaso Evaluación Santillana Ciencias Naturales 5yosiNoch keine Bewertungen
- Guia Séptimo Nutrición en Hongos, AnimalesDokument10 SeitenGuia Séptimo Nutrición en Hongos, Animalestatiana garciaNoch keine Bewertungen
- Aparato Digestivo 2Dokument9 SeitenAparato Digestivo 2Lucelia Ramirez MillaNoch keine Bewertungen
- Antomia Bovino PDFDokument14 SeitenAntomia Bovino PDFHumberto Morales RamirezNoch keine Bewertungen
- Informe Anatomia y Fisiologia Del Sistema DigestivoDokument10 SeitenInforme Anatomia y Fisiologia Del Sistema DigestivoJilmar Æ ZuñigaNoch keine Bewertungen
- Monografia Sistema DigestivoDokument9 SeitenMonografia Sistema DigestivoMaryurit VidaurreNoch keine Bewertungen
- Eda 04-Ficha Informativa-Nutricion-Sistema Digestivo y Circulatorio 11-7-22Dokument7 SeitenEda 04-Ficha Informativa-Nutricion-Sistema Digestivo y Circulatorio 11-7-22Vilma YanguaNoch keine Bewertungen
- El Aparato DigestivoDokument8 SeitenEl Aparato DigestivoMaría Dolores Lisón GarcíaNoch keine Bewertungen
- Sistema DigestivoDokument15 SeitenSistema DigestivofnldkldfNoch keine Bewertungen
- Morfofisiología digestiva monogástricosDokument11 SeitenMorfofisiología digestiva monogástricosjose espitiaNoch keine Bewertungen
- Trabajo de FisiologiaDokument2 SeitenTrabajo de Fisiologiakaren ocampoNoch keine Bewertungen
- Informes Practica 1 2 y 3 Morfifisiologia AnimalDokument27 SeitenInformes Practica 1 2 y 3 Morfifisiologia AnimalEDINSON JAVIER ACOSTA EPIANoch keine Bewertungen
- Sistema digestivo: órganos y funcionesDokument13 SeitenSistema digestivo: órganos y funcionesLuis David Lazo CondoriNoch keine Bewertungen
- Nutrición Heterótrofa 2Dokument4 SeitenNutrición Heterótrofa 2Mauricio Laguna SantaNoch keine Bewertungen
- Sistema digestivo: cuestionario sobre sus partes y funcionesDokument3 SeitenSistema digestivo: cuestionario sobre sus partes y funcionesLourdes Cacha RiosNoch keine Bewertungen
- Nutrición en AnimalesDokument17 SeitenNutrición en AnimalesPaula LozanNoch keine Bewertungen
- Monografia Aparato Digestivo y Aparato UrinarioDokument31 SeitenMonografia Aparato Digestivo y Aparato Urinariorixsy valencia condoriNoch keine Bewertungen
- Triptico Sistema DigestivoDokument3 SeitenTriptico Sistema DigestivoLionel AlvaradoNoch keine Bewertungen
- Taller Nut Rici OnDokument11 SeitenTaller Nut Rici OnPATRICIA DAZANoch keine Bewertungen
- 503 - Biologia - 02Dokument8 Seiten503 - Biologia - 02Angelica BuitragoNoch keine Bewertungen
- Ariadna Mejía Muñoz, Práctica 4, 29-09-2021Dokument8 SeitenAriadna Mejía Muñoz, Práctica 4, 29-09-2021AriadnaMejíaMuñozNoch keine Bewertungen
- Texto DigestivoDokument35 SeitenTexto DigestivoSherly ValenciaNoch keine Bewertungen
- Guia Nutricion en Los Seres VivosDokument4 SeitenGuia Nutricion en Los Seres VivosAlejandra Garzon lesmesNoch keine Bewertungen
- Lección 3 NutriciónDokument5 SeitenLección 3 NutriciónJuan AvilaNoch keine Bewertungen
- Aparato DigestivoDokument17 SeitenAparato DigestivoMaríaFernanda VasquezNoch keine Bewertungen
- Sist Digestivo MonogastricosDokument14 SeitenSist Digestivo MonogastricosCristhian Castillo50% (2)
- Tarea 2 - Morfofisiología de La Digestión en MonogástricosDokument10 SeitenTarea 2 - Morfofisiología de La Digestión en MonogástricosGloria Narvaez100% (1)
- 7° Clases de Sistemas DigestivosDokument7 Seiten7° Clases de Sistemas Digestivosdanilsa reyesNoch keine Bewertungen
- El Proceso Digestivo en Los AnimalesDokument21 SeitenEl Proceso Digestivo en Los AnimalesJHONNoch keine Bewertungen
- Trabajo Biología Materia y Energía 4XDokument24 SeitenTrabajo Biología Materia y Energía 4XJavier Santiago BorghiNoch keine Bewertungen
- Aparato digestivo: función y partes del sistema digestivoDokument2 SeitenAparato digestivo: función y partes del sistema digestivoMaria Alejandra Zamora LopezNoch keine Bewertungen
- V1000Dokument31 SeitenV1000Vicpico PicosoNoch keine Bewertungen
- Regla de Tres SimpleDokument4 SeitenRegla de Tres SimpleBrayhan SánchezNoch keine Bewertungen
- Leccion 1 Nivel 4Dokument6 SeitenLeccion 1 Nivel 4Brayhan SánchezNoch keine Bewertungen
- Diagrama de Proceso Analisis Del ProductoDokument5 SeitenDiagrama de Proceso Analisis Del ProductoBrayhan SánchezNoch keine Bewertungen
- Libro 1Dokument28 SeitenLibro 1Brayhan SánchezNoch keine Bewertungen
- 4.3 Misión y Visión de La CarreraDokument1 Seite4.3 Misión y Visión de La CarreraBrayhan SánchezNoch keine Bewertungen
- Evaluación Cualitativa (Anexo 6)Dokument1 SeiteEvaluación Cualitativa (Anexo 6)Brayhan SánchezNoch keine Bewertungen
- Consulta ControlesDokument14 SeitenConsulta ControlesBrayhan SánchezNoch keine Bewertungen
- UNI-CHIMBORAZO Sílabo de Ingeniería de Métodos IDokument9 SeitenUNI-CHIMBORAZO Sílabo de Ingeniería de Métodos IBrayhan SánchezNoch keine Bewertungen
- Diag H-MDokument6 SeitenDiag H-MBrayhan SánchezNoch keine Bewertungen
- Variador de VelocidadDokument12 SeitenVariador de VelocidadBrayhan SánchezNoch keine Bewertungen
- AsistenciaDokument3 SeitenAsistenciaBrayhan SánchezNoch keine Bewertungen
- Cursograma AnaliticoDokument1 SeiteCursograma AnaliticoBrayhan SánchezNoch keine Bewertungen
- Diagrama de Proceso-Análisis Del HombreDokument4 SeitenDiagrama de Proceso-Análisis Del HombreBrayhan Sánchez0% (2)
- Guía de AudioDokument1 SeiteGuía de AudioBrayhan SánchezNoch keine Bewertungen
- Diagrma Hombre Maquina (Servicios)Dokument2 SeitenDiagrma Hombre Maquina (Servicios)Brayhan SánchezNoch keine Bewertungen
- Encuesta PilotoDokument2 SeitenEncuesta PilotoBrayhan SánchezNoch keine Bewertungen
- Ensayo Del CuidadoDokument3 SeitenEnsayo Del CuidadoBrayhan SánchezNoch keine Bewertungen
- FISIOLOGIA DIGESTIVA EN AVES GRUPO N°04 FibraDokument9 SeitenFISIOLOGIA DIGESTIVA EN AVES GRUPO N°04 FibraBrayhan SánchezNoch keine Bewertungen
- G4-Contabilizacion Peps Metodo PromedioDokument11 SeitenG4-Contabilizacion Peps Metodo PromedioBrayhan SánchezNoch keine Bewertungen
- Toyota SDPDokument7 SeitenToyota SDPBrayhan SánchezNoch keine Bewertungen
- TrabajoDokument3 SeitenTrabajoBrayhan SánchezNoch keine Bewertungen
- Fuerzas ConservativasDokument3 SeitenFuerzas ConservativasBrayhan SánchezNoch keine Bewertungen
- Sumas de RiemannDokument11 SeitenSumas de RiemannBrayhan SánchezNoch keine Bewertungen
- Proyecto SimuladoresDokument12 SeitenProyecto SimuladoresBrayhan SánchezNoch keine Bewertungen
- FISIOLOGIA DIGESTIVA EN AVES GRUPO N°04 FibraDokument9 SeitenFISIOLOGIA DIGESTIVA EN AVES GRUPO N°04 FibraBrayhan SánchezNoch keine Bewertungen
- Consulta 3Dokument5 SeitenConsulta 3Brayhan SánchezNoch keine Bewertungen
- Falsificantes de moneda y palabra en el InfiernoDokument2 SeitenFalsificantes de moneda y palabra en el InfiernonicolleNoch keine Bewertungen
- Que Es Una LaptopDokument3 SeitenQue Es Una LaptopCiencias BásicasNoch keine Bewertungen
- Folklore de La CostaDokument6 SeitenFolklore de La CostaANA ISABEL SOPLIN GARCIANoch keine Bewertungen
- Cuestionarios Corregido Sobre Ecoturismo Con Items Mejorados 2020Dokument6 SeitenCuestionarios Corregido Sobre Ecoturismo Con Items Mejorados 2020Jota RamosNoch keine Bewertungen
- Manual de Capacidad de CarreterasDokument20 SeitenManual de Capacidad de CarreterasXavier Cheme0% (1)
- 01-01 Informacion Sobre Los Fusibles y RelevadoresDokument6 Seiten01-01 Informacion Sobre Los Fusibles y RelevadoresabrahamrojasNoch keine Bewertungen
- Ofertas AndorraDokument10 SeitenOfertas AndorraUP GuadalupeNoch keine Bewertungen
- Literatura Fantastica y Ciencia FicciónDokument13 SeitenLiteratura Fantastica y Ciencia FicciónAkzzel FFNoch keine Bewertungen
- EMEA ThermoPro TP8 PDFDokument2 SeitenEMEA ThermoPro TP8 PDFAlex RogelioNoch keine Bewertungen
- EVSO0629Dokument7 SeitenEVSO0629Editorial El VigiaNoch keine Bewertungen
- Relacion de TiempoDokument5 SeitenRelacion de TiempoEspinoza ErickNoch keine Bewertungen
- Letras Canciones Vol 1 2do Ciclo 2023Dokument5 SeitenLetras Canciones Vol 1 2do Ciclo 2023jugando con elandNoch keine Bewertungen
- Listado de Figuras RetóricasDokument3 SeitenListado de Figuras RetóricasescanquemaNoch keine Bewertungen
- Línea Del Tiempo 10Dokument5 SeitenLínea Del Tiempo 10jesusNoch keine Bewertungen
- Warhammer Aos Rules EsDokument4 SeitenWarhammer Aos Rules EspepeNoch keine Bewertungen
- Estimulacion Cognitiva CuadernilloDokument38 SeitenEstimulacion Cognitiva Cuadernillogabriela nuñezNoch keine Bewertungen
- ClasificacionDeLosInstumentos ENMANUEL PALOMINODokument4 SeitenClasificacionDeLosInstumentos ENMANUEL PALOMINOSnakerubik77 FvoNoch keine Bewertungen
- Juego, dibujo e intervenciones en el análisis infantilDokument19 SeitenJuego, dibujo e intervenciones en el análisis infantilawka_malen100% (1)
- Periféricos No Estandarizados A Través de InterfazDokument9 SeitenPeriféricos No Estandarizados A Través de InterfazYajaira SanzNoch keine Bewertungen
- La Fiesta Es Una Obligación - Completo-2Dokument376 SeitenLa Fiesta Es Una Obligación - Completo-2Quinde SihuarNoch keine Bewertungen
- Trabajo Comentario PlatoDokument1 SeiteTrabajo Comentario Platonai comesañaNoch keine Bewertungen
- RondasInfantilesDokument15 SeitenRondasInfantilesMelissa DE GomezNoch keine Bewertungen
- Descubre Neyba, uno de los pueblos más bonitos de la regiónDokument12 SeitenDescubre Neyba, uno de los pueblos más bonitos de la regiónhdddNoch keine Bewertungen
- Recambios Citroen y PeugeotDokument77 SeitenRecambios Citroen y PeugeotEstela VicenteNoch keine Bewertungen
- Tipos de NarradorDokument9 SeitenTipos de NarradorAri900Noch keine Bewertungen
- Control de Sellos Al DiarioDokument159 SeitenControl de Sellos Al DiarioSite3 VeoliaNoch keine Bewertungen
- Hablo Sobre Mi Rutina Diaria: ConversaciónDokument37 SeitenHablo Sobre Mi Rutina Diaria: ConversaciónJeanNoch keine Bewertungen