Das könnte Ihnen auch gefallen
- ADN (Français) ...Dokument11 SeitenADN (Français) ...Visar NurediniNoch keine Bewertungen
- Les Êtres Vivants Dépendent De La Mécanique QuantiqueVon EverandLes Êtres Vivants Dépendent De La Mécanique QuantiqueNoch keine Bewertungen
- Le hasard au cœur de la cellule: Probabilités, déterminisme, génétiqueVon EverandLe hasard au cœur de la cellule: Probabilités, déterminisme, génétiqueNoch keine Bewertungen
- Qu AntiqueDokument53 SeitenQu AntiqueRayon SoleilNoch keine Bewertungen
- Guide Sur L'énergie ScalaireDokument6 SeitenGuide Sur L'énergie Scalairemichaela doriaNoch keine Bewertungen
- Joël Sternheimer - Le Lieu de La Distinction Sujet-Objet Dans Les Sciences de La Nature.Dokument11 SeitenJoël Sternheimer - Le Lieu de La Distinction Sujet-Objet Dans Les Sciences de La Nature.Michel Blanc100% (1)
- Antenna Zamperini Fran+ºaisDokument5 SeitenAntenna Zamperini Fran+ºaisfreenergiaNoch keine Bewertungen
- TheorieFinalisee09 2016 PDFDokument301 SeitenTheorieFinalisee09 2016 PDFcimpocteNoch keine Bewertungen
- ADN Elisheanportesdutemps Terrenouvelle CA P 311Dokument82 SeitenADN Elisheanportesdutemps Terrenouvelle CA P 311karlaugustt123Noch keine Bewertungen
- Comment Notre Cerveau Produit-Il Des Ondes Scalaires - CCSTIBDokument6 SeitenComment Notre Cerveau Produit-Il Des Ondes Scalaires - CCSTIBSeraphin Miguel100% (1)
- Psyche QuAntiqueDokument6 SeitenPsyche QuAntiqueደጀን ኣባባNoch keine Bewertungen
- Aura ZDokument65 SeitenAura ZkozlhooNoch keine Bewertungen
- Biophoton - La Lumière Est Vivante Elle Communique Avec Toutes Nos CellulesDokument4 SeitenBiophoton - La Lumière Est Vivante Elle Communique Avec Toutes Nos CellulesOrdjounNoch keine Bewertungen
- Bref Récapitulatif KesheDokument18 SeitenBref Récapitulatif KesheAdhi TthanaNoch keine Bewertungen
- Elisheanportesdutemps Terrenouvelle CA P 1240Dokument39 SeitenElisheanportesdutemps Terrenouvelle CA P 1240karlaugustt123Noch keine Bewertungen
- Colette Et Georges Tiret - Survie Et MétamorphosesDokument63 SeitenColette Et Georges Tiret - Survie Et Métamorphosesdigital_dollNoch keine Bewertungen
- Champs ScalaireDokument4 SeitenChamps ScalaireBrigitte NolanNoch keine Bewertungen
- Nexus 67 - Biophysique - Protéodie - La Petite Musique Du Vivant Par Jocelin Morisson (Mars 2010)Dokument8 SeitenNexus 67 - Biophysique - Protéodie - La Petite Musique Du Vivant Par Jocelin Morisson (Mars 2010)ustensilNoch keine Bewertungen
- De l'Univers Physique au Cosmos Métaphysique. L'Intrication Quantique et la Synchronicité de Carl JungVon EverandDe l'Univers Physique au Cosmos Métaphysique. L'Intrication Quantique et la Synchronicité de Carl JungNoch keine Bewertungen
- Étoiles: Les Grands Articles d'UniversalisVon EverandÉtoiles: Les Grands Articles d'UniversalisNoch keine Bewertungen
- Histoire de La Notion de Fréquence SonoreDokument8 SeitenHistoire de La Notion de Fréquence SonoreAlain BoudetNoch keine Bewertungen
- Machine RifeDokument4 SeitenMachine RifePedro Chico MartinNoch keine Bewertungen
- Les Enigmes de la Psychométrie et les Phénomènes de la TélesthésieVon EverandLes Enigmes de la Psychométrie et les Phénomènes de la TélesthésieNoch keine Bewertungen
- Présence: Ovnis, Crop Circles et ExocivilisationsVon EverandPrésence: Ovnis, Crop Circles et ExocivilisationsNoch keine Bewertungen
- L'ADN - Émetteur de Photon Et de Corps de LumièreDokument4 SeitenL'ADN - Émetteur de Photon Et de Corps de Lumièremarcel kefirNoch keine Bewertungen
- Biophotons 001Dokument4 SeitenBiophotons 001jean-louis lavail100% (1)
- Les OrmusDokument8 SeitenLes Ormustfou0076Noch keine Bewertungen
- Les Structures de La MatièreDokument55 SeitenLes Structures de La MatièreAlain Boudet100% (2)
- L'eau en Tant Que Système Rédox (József ORSZÁGH)Dokument15 SeitenL'eau en Tant Que Système Rédox (József ORSZÁGH)GérardNoch keine Bewertungen
- Bousquet Jacqueline - Nature Et Cause Des Maladies PDFDokument12 SeitenBousquet Jacqueline - Nature Et Cause Des Maladies PDFMike Jackson100% (2)
- Article Quantique Psycho Erik PiganiDokument2 SeitenArticle Quantique Psycho Erik PiganiErik PiganiNoch keine Bewertungen
- Bioélectronique Concepts de Base (József ORSZÁGH)Dokument11 SeitenBioélectronique Concepts de Base (József ORSZÁGH)GérardNoch keine Bewertungen
- Nassim Haramein Conference FRDokument116 SeitenNassim Haramein Conference FRMichel ManginNoch keine Bewertungen
- Nexus 45 - Quand La Psychotronique Remplace Les ArmesDokument11 SeitenNexus 45 - Quand La Psychotronique Remplace Les ArmesustensilNoch keine Bewertungen
- A l'heure des synchronicités: Psyché quantique & Expérience perceptive directeVon EverandA l'heure des synchronicités: Psyché quantique & Expérience perceptive directeNoch keine Bewertungen
- 1ère PartieDokument74 Seiten1ère PartiebanzailoicNoch keine Bewertungen
- Pinel ArticleDokument15 SeitenPinel ArticlewebylomeNoch keine Bewertungen
- Radu Cinamar - Decouverte Au BucegiDokument221 SeitenRadu Cinamar - Decouverte Au Bucegiraoul ducoolNoch keine Bewertungen
- Article SternheimerDokument6 SeitenArticle SternheimerJenna Laforge100% (1)
- Basarab Nicolescu, DE LA PHYSIQUE QUANTIQUE AU RÉENCHANTEMENT DU MONDEDokument14 SeitenBasarab Nicolescu, DE LA PHYSIQUE QUANTIQUE AU RÉENCHANTEMENT DU MONDEBasarab Nicolescu100% (3)
- Leadbeater CW Le Plan MentalDokument88 SeitenLeadbeater CW Le Plan MentalArchy AlchilNoch keine Bewertungen
- TP3 Tissus ConjonctifsDokument33 SeitenTP3 Tissus ConjonctifsHadrien De Greef100% (1)
- KISHORI AIRD 2012 Nouvelle Matrice de L UnitEDokument4 SeitenKISHORI AIRD 2012 Nouvelle Matrice de L UnitEyurlungur_frNoch keine Bewertungen
- KESHE TechnologieDokument13 SeitenKESHE TechnologiemcdracheNoch keine Bewertungen
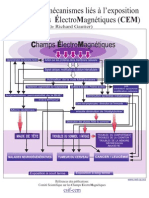
- Schema Mecanismes Interactions Exposition Aux CEMDokument1 SeiteSchema Mecanismes Interactions Exposition Aux CEMpretredutempsNoch keine Bewertungen
- Les phénomènes psychiques occultes: État actuel de la questionVon EverandLes phénomènes psychiques occultes: État actuel de la questionNoch keine Bewertungen
- L'île Des Veilleurs - Le Grand HéritageDokument46 SeitenL'île Des Veilleurs - Le Grand HéritageNawel Bentaïeb100% (2)
- Barbara Hand Clow - The Pleiadian Agenda in FrenchDokument15 SeitenBarbara Hand Clow - The Pleiadian Agenda in FrenchJulien487100% (2)
- ONDES SCALAIRES TeslaDokument10 SeitenONDES SCALAIRES TeslaBESNoch keine Bewertungen
- L'ADN Communique Dans L'universDokument6 SeitenL'ADN Communique Dans L'universGreenLegend100% (1)
- Technologie Keshe en FrançaisDokument11 SeitenTechnologie Keshe en Françaisveronique henry100% (4)
- INTRA TERRE 22 Septembre 2005 Article7025Dokument5 SeitenINTRA TERRE 22 Septembre 2005 Article7025André MANoch keine Bewertungen
- Transition dimensionnelle: Comprendre le passage actuel vécu par l'humanitéVon EverandTransition dimensionnelle: Comprendre le passage actuel vécu par l'humanitéNoch keine Bewertungen
- Rocard InterviewDokument3 SeitenRocard Interviewfredji34Noch keine Bewertungen
- DocmagnetoDokument10 SeitenDocmagnetoBrigitte Nolan100% (1)
- L'Holodynamique Quantique Par Jean-Marc Roeder Et Siegfried de ChaudunDokument20 SeitenL'Holodynamique Quantique Par Jean-Marc Roeder Et Siegfried de ChaudunalexNoch keine Bewertungen
- PURE SANTE-10decouvertesDokument28 SeitenPURE SANTE-10decouvertesjean-loïcNoch keine Bewertungen
- Methylation ADNDokument235 SeitenMethylation ADNjean-loïcNoch keine Bewertungen
- Derrière Le SoleilDokument8 SeitenDerrière Le SoleilHammadi AbidNoch keine Bewertungen
- Dialogues Avec L'ange - Gitta MallaszDokument198 SeitenDialogues Avec L'ange - Gitta MallaszBORIS86% (7)
- Magnetisme Et MoléculesDokument36 SeitenMagnetisme Et Moléculesjean-loïcNoch keine Bewertungen
- ShandarDokument2 SeitenShandarjean-loïcNoch keine Bewertungen
- Yi King PhilastreDokument1.272 SeitenYi King Philastreyiking100% (4)
- Radiesthesie IIDokument28 SeitenRadiesthesie IIgussvii100% (4)
- Philippe Guillemant 9octobre2014Dokument3 SeitenPhilippe Guillemant 9octobre2014jean-loïcNoch keine Bewertungen
- Pendule Egyptien Et Pyramide Ledain PDFDokument69 SeitenPendule Egyptien Et Pyramide Ledain PDFlinden2150100% (7)
- DieuDokument23 SeitenDieujean-loïcNoch keine Bewertungen
- PsychothérapieDokument22 SeitenPsychothérapiekurtkNoch keine Bewertungen
- Petit Guide de La Spiritual 1Dokument40 SeitenPetit Guide de La Spiritual 1Andriamahefamanana Onja TantelyNoch keine Bewertungen
- Electro AcupunctureDokument13 SeitenElectro AcupunctureCarlos MoreiraNoch keine Bewertungen
- La Voix IntérieureDokument8 SeitenLa Voix Intérieurejean-loïcNoch keine Bewertungen
- La Vie Est-Elle Une Succession de Hasards ?Dokument51 SeitenLa Vie Est-Elle Une Succession de Hasards ?Aquarelle100% (2)
- KinesiologiaDokument286 SeitenKinesiologiaManuelNoch keine Bewertungen
- Je Suis Guide Pratique D'éveil Spirituel - Foltzer AntoineDokument80 SeitenJe Suis Guide Pratique D'éveil Spirituel - Foltzer AntoinefghjfghjfghNoch keine Bewertungen
- Le Livre de Vie Des Maitres AscensionnesDokument129 SeitenLe Livre de Vie Des Maitres AscensionnesFabrice Blanc100% (7)
- Electron Immortel - Jean CharonDokument7 SeitenElectron Immortel - Jean Charonjean-loïcNoch keine Bewertungen
- Devère Henri - La Magie Secrète Et Bénéfique de L'huileDokument103 SeitenDevère Henri - La Magie Secrète Et Bénéfique de L'huilejean-loïc100% (2)
- Devenir ChamanDokument94 SeitenDevenir Chamanalterego-33100% (3)
- 194 KabalmacDokument88 Seiten194 Kabalmacهرمس ترجمجيستيس الهندسة المقدسةNoch keine Bewertungen
- Einstein BohmDokument29 SeitenEinstein Bohmjean-loïc100% (1)
- Deunov Peter - Le Livre de La LumièreDokument245 SeitenDeunov Peter - Le Livre de La Lumièrejean-loïcNoch keine Bewertungen
- Divine InterventionDokument8 SeitenDivine Interventionjean-loïcNoch keine Bewertungen
- Champ VitalDokument20 SeitenChamp Vitaljean-loïcNoch keine Bewertungen
- 900 Thoughts Master Peter DeunovDokument87 Seiten900 Thoughts Master Peter DeunovBia De OliveiraNoch keine Bewertungen
- Michel Berçot - Coeur Et ÉnergétiqueDokument396 SeitenMichel Berçot - Coeur Et ÉnergétiqueSheepo Dé la Vega100% (3)
- Remediation CognitiveDokument4 SeitenRemediation CognitiveSteve Innocent EMINI EKONoch keine Bewertungen
- Les-pouvoirs-incroyables-du-cerveau-Amine-Mestari-FrenchPDF.com_Dokument219 SeitenLes-pouvoirs-incroyables-du-cerveau-Amine-Mestari-FrenchPDF.com_HEINNE WITCHOUE100% (2)
- IFPEC Les Fondamentaux de La Psychologie Énergétique FeinsteinDokument60 SeitenIFPEC Les Fondamentaux de La Psychologie Énergétique FeinsteinOlivier BenarrocheNoch keine Bewertungen
- Cours de PsychoDokument6 SeitenCours de PsychoCoby ZehNoch keine Bewertungen
- Animaux CoursDokument8 SeitenAnimaux CoursNahimé YoussefNoch keine Bewertungen
- Ouerchfani s2Dokument14 SeitenOuerchfani s2ryhemchaabeniNoch keine Bewertungen
- Chi Nei Tsang 2Dokument5 SeitenChi Nei Tsang 2ArnoHon100% (1)
- Syndromes TopographiquesDokument30 SeitenSyndromes TopographiquesSafaa MedNoch keine Bewertungen
- 6 MEM PSYCHOMOT 2017 BROUILLIARD-ROUSEAU Ludivine PDFDokument113 Seiten6 MEM PSYCHOMOT 2017 BROUILLIARD-ROUSEAU Ludivine PDFSassiNoch keine Bewertungen
- Révision ST 3e SecDokument62 SeitenRévision ST 3e SecAstride Firdaws BoudaNoch keine Bewertungen
- Dia sn6 SynapsesDokument40 SeitenDia sn6 Synapsesكوردورلي الحاج100% (2)
- Burkina 2014 SVT Serie D 1er Tour Sujet 2 RempDokument4 SeitenBurkina 2014 SVT Serie D 1er Tour Sujet 2 RempbigtoureNoch keine Bewertungen
- Bilans Neuropsychologiques - Mme LEISSING-DESPREZDokument58 SeitenBilans Neuropsychologiques - Mme LEISSING-DESPREZEccoNoch keine Bewertungen
- Nago Livre 1Dokument35 SeitenNago Livre 1Jimmythrio Louis-jeuneNoch keine Bewertungen
- Je Suis Epuise.e Cathy AssenheimDokument224 SeitenJe Suis Epuise.e Cathy AssenheimHichem Hilal100% (1)
- 20 - Favoriser Le Developpement Du CerveauDokument20 Seiten20 - Favoriser Le Developpement Du CerveauzoumNoch keine Bewertungen
- 1-Résumé Lydia Tadjadit Part1 PDFDokument41 Seiten1-Résumé Lydia Tadjadit Part1 PDFSaibiNoch keine Bewertungen
- AVC Et Mouvements Anormaux PDFDokument31 SeitenAVC Et Mouvements Anormaux PDFEmir Salim DjaougaNoch keine Bewertungen
- Hanish Principe AlimentaireDokument152 SeitenHanish Principe AlimentairePhilippe BeugniezNoch keine Bewertungen
- Processus GrabavoiDokument65 SeitenProcessus GrabavoiSerenity Walsh100% (14)