Das könnte Ihnen auch gefallen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- TypeFinderReport ENFPDokument10 SeitenTypeFinderReport ENFPBassant AdelNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Distributing Business Partner Master Data From SAP CRMDokument28 SeitenDistributing Business Partner Master Data From SAP CRMJarko RozemondNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Effect of Intensive Health Education On Adherence To Treatment in Sputum Positive Pulmonary Tuberculosis PatientsDokument6 SeitenEffect of Intensive Health Education On Adherence To Treatment in Sputum Positive Pulmonary Tuberculosis PatientspocutindahNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Unidad 12 (libro-PowerPoint)Dokument5 SeitenUnidad 12 (libro-PowerPoint)Franklin Suarez.HNoch keine Bewertungen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Screening: of Litsea Salicifolia (Dighloti) As A Mosquito RepellentDokument20 SeitenScreening: of Litsea Salicifolia (Dighloti) As A Mosquito RepellentMarmish DebbarmaNoch keine Bewertungen
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Ib Physics SL - Unit 4 ReviewDokument46 SeitenIb Physics SL - Unit 4 ReviewMax HudgenesNoch keine Bewertungen
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- 2nd Form Sequence of WorkDokument7 Seiten2nd Form Sequence of WorkEustace DavorenNoch keine Bewertungen
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Em 1.4 RMDokument18 SeitenEm 1.4 RMMangam RajkumarNoch keine Bewertungen
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Sumit Kataruka Class: Bcom 3 Yr Room No. 24 Roll No. 611 Guide: Prof. Vijay Anand SahDokument20 SeitenSumit Kataruka Class: Bcom 3 Yr Room No. 24 Roll No. 611 Guide: Prof. Vijay Anand SahCricket KheloNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Invitation 2023Dokument10 SeitenInvitation 2023Joanna Marie Cruz FelipeNoch keine Bewertungen
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Reported SpeechDokument2 SeitenReported SpeechmayerlyNoch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- An Analysis of The PoemDokument2 SeitenAn Analysis of The PoemDayanand Gowda Kr100% (2)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- CH Folk Media and HeatlhDokument6 SeitenCH Folk Media and HeatlhRaghavendr KoreNoch keine Bewertungen
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Tan vs. Macapagal, 43 SCRADokument6 SeitenTan vs. Macapagal, 43 SCRANikkaDoriaNoch keine Bewertungen
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- What Is Thesis Plural FormDokument8 SeitenWhat Is Thesis Plural Formtracyjimenezstamford100% (2)
- Site Master S113C, S114C, S331C, S332C, Antenna, Cable and Spectrum AnalyzerDokument95 SeitenSite Master S113C, S114C, S331C, S332C, Antenna, Cable and Spectrum AnalyzerKodhamagulla SudheerNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Sysmex HemostasisDokument11 SeitenSysmex HemostasisElyza L. de GuzmanNoch keine Bewertungen
- Erika Nuti Chrzsoloras Pewri Tou Basileos LogouDokument31 SeitenErika Nuti Chrzsoloras Pewri Tou Basileos Logouvizavi21Noch keine Bewertungen
- Level 5 - LFH 6-10 SepDokument14 SeitenLevel 5 - LFH 6-10 SepJanna GunioNoch keine Bewertungen
- Eet 223 (1) Analog Electronics JagjeetDokument79 SeitenEet 223 (1) Analog Electronics JagjeetMahima ArrawatiaNoch keine Bewertungen
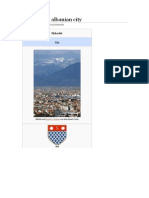
- Shkodër An Albanian CityDokument16 SeitenShkodër An Albanian CityXINKIANGNoch keine Bewertungen
- "We Like": Rohit Kiran KeluskarDokument43 Seiten"We Like": Rohit Kiran Keluskarrohit keluskarNoch keine Bewertungen
- Saxonville CaseDokument2 SeitenSaxonville Casel103m393Noch keine Bewertungen
- Literature Review On Catfish ProductionDokument5 SeitenLiterature Review On Catfish Productionafmzyodduapftb100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- DigoxinDokument18 SeitenDigoxinApril Mergelle LapuzNoch keine Bewertungen
- Designing A Peace Building InfrastructureDokument253 SeitenDesigning A Peace Building InfrastructureAditya SinghNoch keine Bewertungen
- Roofing Shingles in KeralaDokument13 SeitenRoofing Shingles in KeralaCertainteed Roofing tilesNoch keine Bewertungen
- Administrator's Guide: SeriesDokument64 SeitenAdministrator's Guide: SeriesSunny SaahilNoch keine Bewertungen
- Mirza HRM ProjectDokument44 SeitenMirza HRM Projectsameer82786100% (1)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- K.M Nanavati v. State of MaharashtraDokument6 SeitenK.M Nanavati v. State of MaharashtraPushpank PandeyNoch keine Bewertungen
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)