Das könnte Ihnen auch gefallen
- Bioquímica Informe N°8 y 9 - Cinética EnzimáticaDokument22 SeitenBioquímica Informe N°8 y 9 - Cinética EnzimáticaMestizo Sinalefa50% (6)
- Marco Teorico Actividad EnzimaticaDokument3 SeitenMarco Teorico Actividad EnzimaticaLeandro LópezNoch keine Bewertungen
- La Ciencia Del Metabolismo: Metabolismo sin rodeosVon EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosBewertung: 5 von 5 Sternen5/5 (1)
- Cinética EnzimáticaDokument41 SeitenCinética EnzimáticaClaudia HernándezNoch keine Bewertungen
- Enzimología clínica: marcadores, métodos y factoresDokument85 SeitenEnzimología clínica: marcadores, métodos y factoresRuben Larico LopezNoch keine Bewertungen
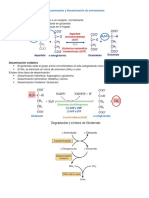
- Transaminación y Desaminación de AminoácidosDokument4 SeitenTransaminación y Desaminación de Aminoácidos宇董Noch keine Bewertungen
- Enzimología ClínicaDokument25 SeitenEnzimología ClínicaJhumira Perez DelaoNoch keine Bewertungen
- Cinética enzimática de la invertasaDokument8 SeitenCinética enzimática de la invertasaAndy Phy100% (1)
- Intoxicacion Por FármacosDokument34 SeitenIntoxicacion Por FármacosLeonardo100% (1)
- Miastenia Gravis HSHSHSHDokument34 SeitenMiastenia Gravis HSHSHSHJose VacaNoch keine Bewertungen
- Gases Arteriales-InterpretaciónDokument89 SeitenGases Arteriales-InterpretaciónRaKeLiTaZ0% (1)
- Ev4 - Indicadores Enzimaticos No FuncionalesDokument9 SeitenEv4 - Indicadores Enzimaticos No FuncionalesGustavo EstradaNoch keine Bewertungen
- Enzimas y Pancreático ####Dokument62 SeitenEnzimas y Pancreático ####7z6v7d7dhyNoch keine Bewertungen
- Discusion 3Dokument7 SeitenDiscusion 3José LuisNoch keine Bewertungen
- Propiedades EnzimasDokument7 SeitenPropiedades EnzimasRené Belisario Carranza56% (9)
- Enzimología ClinicaDokument21 SeitenEnzimología ClinicaEdsonVictorPalominoNoch keine Bewertungen
- Guía Actividad EnzimáticaDokument15 SeitenGuía Actividad EnzimáticaDaniela AdarmeNoch keine Bewertungen
- Enzimas de Intéres ClínicoDokument54 SeitenEnzimas de Intéres ClínicoMario Gonzalez Olguin100% (2)
- Practica 5 Determinacion de EnzimaDokument16 SeitenPractica 5 Determinacion de Enzimaliz92070% (1)
- Resumen Qca Uabp 4Dokument6 SeitenResumen Qca Uabp 4Osvaldo HerraezNoch keine Bewertungen
- 2-Localizacion Celular y Cinetica EnzimaticaDokument28 Seiten2-Localizacion Celular y Cinetica EnzimaticaJC XmanNoch keine Bewertungen
- Seminario de EnzimologiaDokument55 SeitenSeminario de EnzimologiahikarujjNoch keine Bewertungen
- Amilasa y LipasaDokument13 SeitenAmilasa y LipasaKenia FragosoNoch keine Bewertungen
- Enzimología clínica: definición, tipos de enzimas y su aplicación en el diagnóstico médicoDokument5 SeitenEnzimología clínica: definición, tipos de enzimas y su aplicación en el diagnóstico médicoAndreaBenitezNoch keine Bewertungen
- Bioquimica y Biologia MolecularDokument41 SeitenBioquimica y Biologia MolecularRaysa Yadira Alberca AtaramaNoch keine Bewertungen
- Enzimas y Clasificacion de EnzimasDokument20 SeitenEnzimas y Clasificacion de EnzimasAlejandro Gomez DiazNoch keine Bewertungen
- Actividad Enzimatica GOTDokument6 SeitenActividad Enzimatica GOTKEVIN DANIEL ORTEGA FRANCONoch keine Bewertungen
- Enzimologia Clínica 2022Dokument75 SeitenEnzimologia Clínica 2022Fanny ChavezNoch keine Bewertungen
- Enzimas Biologicas de Interes ClinicoDokument8 SeitenEnzimas Biologicas de Interes ClinicoMaria Fernanda PalmaNoch keine Bewertungen
- Estudio cinético de la ureasaDokument23 SeitenEstudio cinético de la ureasalionz1010Noch keine Bewertungen
- AminoacidosDokument91 SeitenAminoacidosLeidi BarreraNoch keine Bewertungen
- Transam in As AsDokument5 SeitenTransam in As AssteffyNoch keine Bewertungen
- 11 FatimaDokument9 Seiten11 FatimaFátima Barrios Wong50% (2)
- Perfil de Lipidos 1Dokument28 SeitenPerfil de Lipidos 1GonzaloNoch keine Bewertungen
- Discusion 3 Bioca IDokument12 SeitenDiscusion 3 Bioca IOmar VásquezNoch keine Bewertungen
- Cinetica EnzimaticaDokument13 SeitenCinetica EnzimaticaLaura CastilloNoch keine Bewertungen
- EnzimologiaDokument14 SeitenEnzimologiadaniyamiletteNoch keine Bewertungen
- En ZimasDokument15 SeitenEn ZimasCarlos CanchisNoch keine Bewertungen
- Bioquimica Informe Sem 1Dokument18 SeitenBioquimica Informe Sem 1Vivian Sotomayor FloresNoch keine Bewertungen
- Discusión 2.0 PDFDokument33 SeitenDiscusión 2.0 PDFSaul ArguetaNoch keine Bewertungen
- Resumen de Bioquimic1manuelDokument5 SeitenResumen de Bioquimic1manuelManuel Angel Fernandez CriolloNoch keine Bewertungen
- En ZimasDokument17 SeitenEn ZimasSERGIO MARTINEZ ROMERONoch keine Bewertungen
- En ZimasDokument17 SeitenEn ZimasSERGIO MARTINEZ ROMERONoch keine Bewertungen
- Bioquímica I (laboratorio 2: EnzimasDokument7 SeitenBioquímica I (laboratorio 2: EnzimasKenia GrijalvaNoch keine Bewertungen
- Discusion de Grupo N 3Dokument14 SeitenDiscusion de Grupo N 3RubenPerez100% (4)
- Práctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Dokument11 SeitenPráctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Angel Gustavo HernandezNoch keine Bewertungen
- UntitledDokument40 SeitenUntitledJORDAN VILLEGAS OSORESNoch keine Bewertungen
- Practica de Laboratorio 4 - TrasminasasDokument6 SeitenPractica de Laboratorio 4 - TrasminasasFiorella DiosesNoch keine Bewertungen
- ENZIMIOLOGÍA CLINICA ENSAYO BioquimicaDokument5 SeitenENZIMIOLOGÍA CLINICA ENSAYO BioquimicaMauren Molina RiveraNoch keine Bewertungen
- Práctica N4Dokument19 SeitenPráctica N4Jesus SiesquenNoch keine Bewertungen
- Enzimologia ClinicaDokument31 SeitenEnzimologia ClinicaStefanny OrtegaNoch keine Bewertungen
- Ensayo Enzimología Clínica Universidad CartagenaDokument5 SeitenEnsayo Enzimología Clínica Universidad CartagenaEstefani ChavezNoch keine Bewertungen
- Actividad EnzimáticaDokument11 SeitenActividad EnzimáticaEnmanuel Samudio100% (1)
- ENZIMASDokument41 SeitenENZIMASnicolehurtadosNoch keine Bewertungen
- Enzimas y Actividad EnzimaticaDokument28 SeitenEnzimas y Actividad EnzimaticaJuan Carlos AnicamaNoch keine Bewertungen
- Propiedades de Los EnzimasDokument3 SeitenPropiedades de Los EnzimasKevin Jose PatiñoNoch keine Bewertungen
- Tema 4-2Dokument5 SeitenTema 4-2teresarodriguezgallegosNoch keine Bewertungen
- Enzimología ClínicaDokument11 SeitenEnzimología ClínicaLlencis ColónNoch keine Bewertungen
- Tema 6 Analisis BioquimicoDokument5 SeitenTema 6 Analisis BioquimicoSieliaNoch keine Bewertungen
- Actividad Enzimatica y PHDokument12 SeitenActividad Enzimatica y PHxiomaraNoch keine Bewertungen
- Bbioquimica de Los Alimentos Enzimas en Panaderia y ConfiteriaDokument37 SeitenBbioquimica de Los Alimentos Enzimas en Panaderia y ConfiteriaJerson David Rubín TorresNoch keine Bewertungen
- Enzimas bioquímicas: definición, propiedades y aplicaciones clínicasDokument4 SeitenEnzimas bioquímicas: definición, propiedades y aplicaciones clínicasRosmar MejiasNoch keine Bewertungen
- Cito Genetic ADokument10 SeitenCito Genetic A宇董Noch keine Bewertungen
- PorfirinaDokument1 SeitePorfirina宇董Noch keine Bewertungen
- Inmunidad AntitumoralDokument8 SeitenInmunidad Antitumoral宇董Noch keine Bewertungen
- Fisiología de La Transmisión Colinérgica y AdrenérgicaDokument6 SeitenFisiología de La Transmisión Colinérgica y Adrenérgica宇董Noch keine Bewertungen
- Los TelómerosDokument3 SeitenLos Telómeros宇董Noch keine Bewertungen
- INFORME BIOQUIMICA (Actividad Enzimatica)Dokument5 SeitenINFORME BIOQUIMICA (Actividad Enzimatica)Diego JavierNoch keine Bewertungen
- Guion Final MyN Oct 2010Dokument15 SeitenGuion Final MyN Oct 2010宇董Noch keine Bewertungen
- Larrain JDokument47 SeitenLarrain J宇董Noch keine Bewertungen
- Digestión y transporte de proteínas enDokument7 SeitenDigestión y transporte de proteínas en宇董Noch keine Bewertungen
- 6 EdemaDokument3 Seiten6 EdemaLaura GuardiaNoch keine Bewertungen
- Bio Medic in A 01Dokument104 SeitenBio Medic in A 01pmedel@gmail.comNoch keine Bewertungen
- HipertiroidismoDokument5 SeitenHipertiroidismoEdixon LealNoch keine Bewertungen
- Clase VII - 2022 - Inmunología e HipersensibilidadDokument28 SeitenClase VII - 2022 - Inmunología e HipersensibilidadMaite RodriguezNoch keine Bewertungen
- Diagnóstico de Laboratorio de Anticuerpos Antifosfolípidos / 2016Dokument9 SeitenDiagnóstico de Laboratorio de Anticuerpos Antifosfolípidos / 2016Ari EcheverríaNoch keine Bewertungen
- Abp Patologias Humanas Con Alteraciones en El Citoesqueleto TerminadoDokument8 SeitenAbp Patologias Humanas Con Alteraciones en El Citoesqueleto TerminadoAnonymous WbKbQ78Noch keine Bewertungen
- Examen Parcial Anat. 1 2021-1Dokument6 SeitenExamen Parcial Anat. 1 2021-1FERNANDA CLAUDETTE VILLA LAZARONoch keine Bewertungen
- ACP: Caso de agnosia visuoespacial progresivaDokument15 SeitenACP: Caso de agnosia visuoespacial progresivaLina Marcela Marin SotoNoch keine Bewertungen
- 20 Enfermedades Del Tracto DigestivoDokument19 Seiten20 Enfermedades Del Tracto DigestivoSteven MorenoNoch keine Bewertungen
- ANESTESIA I - NotasDokument51 SeitenANESTESIA I - NotasManuela Arcila OrtizNoch keine Bewertungen
- Ram Clasificacion PDFDokument46 SeitenRam Clasificacion PDFDanny Daniel Romero CcasaniNoch keine Bewertungen
- Algunos Tipos de Láser y Su Utilización en MedicinaDokument2 SeitenAlgunos Tipos de Láser y Su Utilización en Medicinaandres26_8Noch keine Bewertungen
- UrticariaDokument30 SeitenUrticariaAlejandro CovarrubiasNoch keine Bewertungen
- Glomerulonefritis en NiñosDokument52 SeitenGlomerulonefritis en NiñosMon BNoch keine Bewertungen
- R-2 T-22 Semiología de Los Genitales Masculinos y FemeninosDokument15 SeitenR-2 T-22 Semiología de Los Genitales Masculinos y FemeninosvaniaNoch keine Bewertungen
- Hipertensión CorregidoDokument61 SeitenHipertensión CorregidoGustavo GaleanoNoch keine Bewertungen
- Clase 6 AntiparkinsonianosDokument30 SeitenClase 6 AntiparkinsonianosNicky BurgosNoch keine Bewertungen
- Mapa - Insuficiencia Cardiaca - Nayeli PintoDokument1 SeiteMapa - Insuficiencia Cardiaca - Nayeli PintoClarita GuerreroNoch keine Bewertungen
- Eritema Nodoso en PediatriaDokument17 SeitenEritema Nodoso en PediatriaMonica Silva BerruzNoch keine Bewertungen
- Les 2Dokument63 SeitenLes 2Tesis ShaioNoch keine Bewertungen
- Resumen PediatriaDokument51 SeitenResumen PediatriaGabriela Szita CeroniNoch keine Bewertungen
- Celulitis (Inflamación) - Wikipedia, La Enciclopedia LibreDokument25 SeitenCelulitis (Inflamación) - Wikipedia, La Enciclopedia LibrejosealopezrNoch keine Bewertungen
- Cirrosis Hépatica DeberDokument18 SeitenCirrosis Hépatica DeberlourdesNoch keine Bewertungen
- 200 años de Independencia: Caso de Leucemia Mieloide CrónicaDokument7 Seiten200 años de Independencia: Caso de Leucemia Mieloide CrónicaFranco IslaNoch keine Bewertungen
- OligohidramniosDokument3 SeitenOligohidramniosAndreha MartínezNoch keine Bewertungen
- Tumores IntracranealesDokument14 SeitenTumores IntracranealesHector BlueNoch keine Bewertungen
- Funcionalismo LeucocitarioDokument54 SeitenFuncionalismo LeucocitarioIsela MendozaNoch keine Bewertungen