Das könnte Ihnen auch gefallen
- Tarea C.Exactas Semana 2Dokument6 SeitenTarea C.Exactas Semana 2Roberto QuirogaNoch keine Bewertungen
- Tercero Hdengue 5 Mandar A Fisicoquimica TambienDokument5 SeitenTercero Hdengue 5 Mandar A Fisicoquimica TambienRoberto QuirogaNoch keine Bewertungen
- Dengue PDFDokument58 SeitenDengue PDFmicaelaNoch keine Bewertungen
- Tercero Hdengue 5 Mandar A Fisicoquimica TambienDokument5 SeitenTercero Hdengue 5 Mandar A Fisicoquimica TambienRoberto QuirogaNoch keine Bewertungen
- Primero 6 Las Células Eucariotas y ProcariotasDokument3 SeitenPrimero 6 Las Células Eucariotas y ProcariotasRoberto QuirogaNoch keine Bewertungen
- Dengue PDFDokument58 SeitenDengue PDFmicaelaNoch keine Bewertungen
- Tercero Genética 6Dokument1 SeiteTercero Genética 6Roberto QuirogaNoch keine Bewertungen
- Primero 6 Las Células Eucariotas y ProcariotasDokument3 SeitenPrimero 6 Las Células Eucariotas y ProcariotasRoberto QuirogaNoch keine Bewertungen
- Primero 1 El Origen de La Vida PDFDokument1 SeitePrimero 1 El Origen de La Vida PDFRoberto QuirogaNoch keine Bewertungen
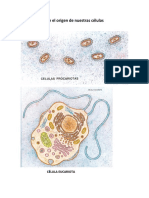
- Primero 4 El Origen de Nuestras CélulasDokument3 SeitenPrimero 4 El Origen de Nuestras CélulasRoberto QuirogaNoch keine Bewertungen
- Tercero Genética 6Dokument1 SeiteTercero Genética 6Roberto QuirogaNoch keine Bewertungen
- Primero 2 El Origen de La Vida PDFDokument3 SeitenPrimero 2 El Origen de La Vida PDFRoberto QuirogaNoch keine Bewertungen
- Primero 5 La Célula Eucariota Estructura y FunciónDokument5 SeitenPrimero 5 La Célula Eucariota Estructura y FunciónRoberto QuirogaNoch keine Bewertungen
- Biología Herencia 3º1Dokument2 SeitenBiología Herencia 3º1Roberto QuirogaNoch keine Bewertungen
- Primero 3 Caracteristicas de Los Seres VivosDokument2 SeitenPrimero 3 Caracteristicas de Los Seres VivosRoberto QuirogaNoch keine Bewertungen
- MocaseDokument30 SeitenMocaseRoberto QuirogaNoch keine Bewertungen
- Primero 2 El Origen de La Vida PDFDokument3 SeitenPrimero 2 El Origen de La Vida PDFRoberto QuirogaNoch keine Bewertungen
- 3 Trabajo Geometría SegundoDokument2 Seiten3 Trabajo Geometría SegundoRoberto QuirogaNoch keine Bewertungen
- Evacuación en CamaronesDokument5 SeitenEvacuación en CamaronesRoberto QuirogaNoch keine Bewertungen
- 2 Trabajo Geometría SegundoDokument2 Seiten2 Trabajo Geometría SegundoRoberto QuirogaNoch keine Bewertungen
- 1 Trabajo Geometría Segundo PDFDokument1 Seite1 Trabajo Geometría Segundo PDFRoberto QuirogaNoch keine Bewertungen
- VoucherDokument2 SeitenVoucherRoberto QuirogaNoch keine Bewertungen
- Planificación Digestivo QuirogaDokument16 SeitenPlanificación Digestivo QuirogaRoberto QuirogaNoch keine Bewertungen
- Movimiento CampesinoDokument8 SeitenMovimiento CampesinoRoberto QuirogaNoch keine Bewertungen
- HIV y VaricelaDokument4 SeitenHIV y VaricelaRoberto QuirogaNoch keine Bewertungen
- Planificación Digestivo QuirogaDokument16 SeitenPlanificación Digestivo QuirogaRoberto QuirogaNoch keine Bewertungen
- Estatuto DocenteDokument7 SeitenEstatuto DocenteRoberto QuirogaNoch keine Bewertungen
- DidacticaDokument1 SeiteDidacticaRoberto QuirogaNoch keine Bewertungen
- LAFIOSCA Politica Unidad 2Dokument130 SeitenLAFIOSCA Politica Unidad 2Roberto QuirogaNoch keine Bewertungen
- Amniotas IDokument14 SeitenAmniotas IRoberto QuirogaNoch keine Bewertungen
- Reporte Escrito: Identificación de Infección Activa Del PatógenoDokument11 SeitenReporte Escrito: Identificación de Infección Activa Del PatógenoJuan Carlos Banda FonsecaNoch keine Bewertungen
- Los Virus (Apuntes)Dokument8 SeitenLos Virus (Apuntes)Ezequiel GuardiaNoch keine Bewertungen
- Deteccion de Anticuerpos Anti Hiv IntroducciónDokument5 SeitenDeteccion de Anticuerpos Anti Hiv IntroducciónRosario ParesNoch keine Bewertungen
- 3.01.17 RabiesDokument38 Seiten3.01.17 RabiesChristhell SanchezNoch keine Bewertungen
- Replicación ViralDokument11 SeitenReplicación ViralSara AlfaroNoch keine Bewertungen
- Del ADN a la proteína: Cómo leen las células el genomaDokument22 SeitenDel ADN a la proteína: Cómo leen las células el genomaAnarcos Andres Canivilo VargasNoch keine Bewertungen
- Banda 2Dokument4 SeitenBanda 2Albert Pinto Tapia CevicheNoch keine Bewertungen
- Microbiología veterinaria: Taxonomía de virus ARNDokument34 SeitenMicrobiología veterinaria: Taxonomía de virus ARNAnonymous s1pg0ks7Noch keine Bewertungen
- Replicación Viral y Ciclos de Multiplicación de Los Virus PDFDokument4 SeitenReplicación Viral y Ciclos de Multiplicación de Los Virus PDFAndy GarciaNoch keine Bewertungen
- Unidad 7: Técnicas de PCRDokument9 SeitenUnidad 7: Técnicas de PCRMary Cruz Collahuazo FloresNoch keine Bewertungen
- Tarea 2.1Dokument3 SeitenTarea 2.1Lydia Gonzalez RodriguezNoch keine Bewertungen
- Sint ProtDokument40 SeitenSint ProtFlorencia BarberisNoch keine Bewertungen
- Complejo TorchDokument43 SeitenComplejo TorchChristian ZapaNoch keine Bewertungen
- Retroviridae familia virus ARNDokument41 SeitenRetroviridae familia virus ARNMaria Alejandra Vargas QuinteroNoch keine Bewertungen
- Exposición Sobre El VIHDokument4 SeitenExposición Sobre El VIHLionel Andrés Messi CuccittiniNoch keine Bewertungen
- Plantas Con Prop Anti-ViralesDokument5 SeitenPlantas Con Prop Anti-ViralesMilagros Aguilar RamosNoch keine Bewertungen
- Generalidades de La Transcripción en EucariotasDokument9 SeitenGeneralidades de La Transcripción en EucariotasClayton JairoNoch keine Bewertungen
- Proyecto - Segundo Parcial - Aplicaciones MatemáticasDokument11 SeitenProyecto - Segundo Parcial - Aplicaciones MatemáticasJorgeLuisFR725% (4)
- Genetica Molecular PDFDokument25 SeitenGenetica Molecular PDFAitana JGNoch keine Bewertungen
- Clasificación de BaltimoreDokument3 SeitenClasificación de BaltimoreadriponsNoch keine Bewertungen
- Wuolah Free TEMA 14 Microbiologia y BiotecnologiaDokument24 SeitenWuolah Free TEMA 14 Microbiologia y BiotecnologiamariaNoch keine Bewertungen
- VIROLOGIADokument21 SeitenVIROLOGIACristian David Adarve MarinNoch keine Bewertungen
- RT PCRDokument4 SeitenRT PCRMaria Del Mar FernándezNoch keine Bewertungen
- Biologia, Inmunología IIDokument32 SeitenBiologia, Inmunología IILinda0% (1)
- Síntesis de cDNADokument26 SeitenSíntesis de cDNALidiaAlejandraRiveraPonceNoch keine Bewertungen
- Trabajo Del SidaDokument15 SeitenTrabajo Del SidaALBERTO266Noch keine Bewertungen
- Enfermedades Tratadas Con Terapia GenicaDokument10 SeitenEnfermedades Tratadas Con Terapia GenicaFabiola Chaparro ReyesNoch keine Bewertungen
- Targa. Seminario de Farmacología.Dokument15 SeitenTarga. Seminario de Farmacología.Jc CastroNoch keine Bewertungen
- Deteccion Vih Western BlotDokument18 SeitenDeteccion Vih Western BlotAlvaro Alejandro Lopez SequeirosNoch keine Bewertungen
- Clase N°2Dokument44 SeitenClase N°2Scarlett AravenaNoch keine Bewertungen