Das könnte Ihnen auch gefallen
- Ii Taller Vect 2020 1Dokument1 SeiteIi Taller Vect 2020 1Bermúdez G ManuelNoch keine Bewertungen
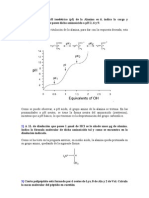
- Taller Estadistica - ResueltoDokument6 SeitenTaller Estadistica - ResueltoTONY NAVARRO NAVARRONoch keine Bewertungen
- Tony Navarro Navarro 2017211035 - Grupo #1Dokument4 SeitenTony Navarro Navarro 2017211035 - Grupo #1TONY NAVARRO NAVARRONoch keine Bewertungen
- Ii Taller Vect 2020 1Dokument1 SeiteIi Taller Vect 2020 1Bermúdez G ManuelNoch keine Bewertungen
- Examen de Resistencia de MaterialesDokument1 SeiteExamen de Resistencia de MaterialesTONY NAVARRO NAVARRONoch keine Bewertungen
- Taller de ResistenciaDokument3 SeitenTaller de ResistenciaTONY NAVARRO NAVARRONoch keine Bewertungen
- Examen de Resistencia de Materiales.2Dokument1 SeiteExamen de Resistencia de Materiales.2TONY NAVARRO NAVARRONoch keine Bewertungen
- TRABAJO CalculoDokument7 SeitenTRABAJO CalculoTONY NAVARRO NAVARRONoch keine Bewertungen
- Examen de Resistencia de Materiales.4Dokument1 SeiteExamen de Resistencia de Materiales.4TONY NAVARRO NAVARRONoch keine Bewertungen
- Registro Observacion Al Desarrollo Mes Septiembre 2020Dokument6 SeitenRegistro Observacion Al Desarrollo Mes Septiembre 2020TONY NAVARRO NAVARRONoch keine Bewertungen
- Examen de Resistencia de Materiales.3Dokument1 SeiteExamen de Resistencia de Materiales.3TONY NAVARRO NAVARRONoch keine Bewertungen
- Taller de Topografia 2Dokument2 SeitenTaller de Topografia 2TONY NAVARRO NAVARRONoch keine Bewertungen
- HUAWEI Y6II Guia de Inicio Rapido CAM-L23 02 Español&English PDFDokument52 SeitenHUAWEI Y6II Guia de Inicio Rapido CAM-L23 02 Español&English PDFTONY NAVARRO NAVARRONoch keine Bewertungen
- Registro Observacion Al Desarrollo Mes Septiembre 2020Dokument6 SeitenRegistro Observacion Al Desarrollo Mes Septiembre 2020TONY NAVARRO NAVARRONoch keine Bewertungen
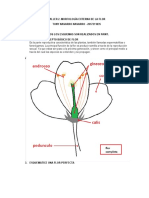
- Taller 2. FlorDokument4 SeitenTaller 2. FlorTONY NAVARRO NAVARRONoch keine Bewertungen
- LECCIÓN 2 Abstracción y Modelamiento CartográficoDokument28 SeitenLECCIÓN 2 Abstracción y Modelamiento CartográficoMake MenroNoch keine Bewertungen
- Historia de La Quimica OrganicaDokument2 SeitenHistoria de La Quimica OrganicaTONY NAVARRO NAVARRONoch keine Bewertungen
- Mecanica Taller UnificadoDokument4 SeitenMecanica Taller UnificadoTONY NAVARRO NAVARRONoch keine Bewertungen
- Morfologia y Anatomia de La RaizDokument5 SeitenMorfologia y Anatomia de La RaizTONY NAVARRO NAVARRONoch keine Bewertungen
- Mecanica Taller UnificadoDokument4 SeitenMecanica Taller UnificadoTONY NAVARRO NAVARRONoch keine Bewertungen
- TopoDokument6 SeitenTopoTONY NAVARRO NAVARRONoch keine Bewertungen
- Programa y Practica de Probabilidad y Estadistica 2012 1Dokument2 SeitenPrograma y Practica de Probabilidad y Estadistica 2012 1TONY NAVARRO NAVARRONoch keine Bewertungen
- CV 39Dokument2 SeitenCV 39MarielenaPerezNoch keine Bewertungen
- FihdkDokument9 SeitenFihdkTONY NAVARRO NAVARRONoch keine Bewertungen
- Programa y Practica de Probabilidad y Estadistica 2012 1Dokument26 SeitenPrograma y Practica de Probabilidad y Estadistica 2012 1Kevin OlveraNoch keine Bewertungen
- Curriculum vitae profesionalDokument1 SeiteCurriculum vitae profesionalmaisaarteaga100% (1)
- Formato4 1Dokument2 SeitenFormato4 1Jhoel Julca RamosNoch keine Bewertungen
- Miproyectodevida 130930210748 Phpapp01Dokument27 SeitenMiproyectodevida 130930210748 Phpapp01Carlos EscobarNoch keine Bewertungen
- Programa y Practica de Probabilidad y Estadistica 2012 1Dokument26 SeitenPrograma y Practica de Probabilidad y Estadistica 2012 1Kevin OlveraNoch keine Bewertungen
- Antropologia en Colombia Jacke - 1Dokument3 SeitenAntropologia en Colombia Jacke - 1TONY NAVARRO NAVARRONoch keine Bewertungen
- English Melatonine - Ilo.esDokument28 SeitenEnglish Melatonine - Ilo.esLiz OrtegaNoch keine Bewertungen
- ThichodermaDokument14 SeitenThichodermaDarwin GuamanNoch keine Bewertungen
- Actividad - Colaborativa - Grupo - 151009 - 37-BiologiaDokument9 SeitenActividad - Colaborativa - Grupo - 151009 - 37-BiologiaFabian Andres Sanchez HernandezNoch keine Bewertungen
- INTA CRBsAs EEABalcarce Brambilla EC Uso Espectrofometría Absorción AtómicaDokument106 SeitenINTA CRBsAs EEABalcarce Brambilla EC Uso Espectrofometría Absorción AtómicaCuellar Melgar Maira AlejandraNoch keine Bewertungen
- Evaluacion de La CelulaDokument2 SeitenEvaluacion de La CelulaEsmeralda CorredorNoch keine Bewertungen
- 1a DR Curay MB y Termoregulación 2022Dokument86 Seiten1a DR Curay MB y Termoregulación 2022CIELO CAROLINA DEL ROSARIO PE�ANoch keine Bewertungen
- Actividad 2BDokument3 SeitenActividad 2BVanquelinNoch keine Bewertungen
- Biocel Lab PDFDokument12 SeitenBiocel Lab PDFCamila FaríasNoch keine Bewertungen
- Conf #5 - Los Polisacaridos - BIDokument25 SeitenConf #5 - Los Polisacaridos - BIYosaira GuerreroNoch keine Bewertungen
- Bioquímica Unidad I Introducción y GeneralidadesDokument16 SeitenBioquímica Unidad I Introducción y GeneralidadesDiana KristinaNoch keine Bewertungen
- Crucigrama La CélulaDokument2 SeitenCrucigrama La CélulaAngel Ruíz GarcíaNoch keine Bewertungen
- Síntesis Proteica: Katherine Sánchez Santamaría Fonoaudióloga Un MSC en Fisiología UnDokument17 SeitenSíntesis Proteica: Katherine Sánchez Santamaría Fonoaudióloga Un MSC en Fisiología UnValentina Orozco AgudeloNoch keine Bewertungen
- Catabolismo de Carbohidratos PDFDokument23 SeitenCatabolismo de Carbohidratos PDFZrtPaoNoch keine Bewertungen
- Informe de BiologiaDokument4 SeitenInforme de Biologiatokio den verNoch keine Bewertungen
- Bioquimicas PresentacionDokument43 SeitenBioquimicas PresentacionJ Armando BritoNoch keine Bewertungen
- Practica Extracción de ADN 3 - 220922 - 230954Dokument7 SeitenPractica Extracción de ADN 3 - 220922 - 230954Jimmy Alexander Marinero LemusNoch keine Bewertungen
- Oxidación de Ácidos Grasos y Cetogénesis PDFDokument37 SeitenOxidación de Ácidos Grasos y Cetogénesis PDFalymbt67% (3)
- Beta-Oxidacion de Ácidos GrasosDokument12 SeitenBeta-Oxidacion de Ácidos GrasosalezNoch keine Bewertungen
- HexocinasaDokument10 SeitenHexocinasamisaedicitaNoch keine Bewertungen
- Consenso Sobre Las Grasas y Aceites 2015 FESNAD PDFDokument80 SeitenConsenso Sobre Las Grasas y Aceites 2015 FESNAD PDFAdrianNoch keine Bewertungen
- 100% Online - Curso Endocrino - Farmacología y Suplementación DeportivaDokument16 Seiten100% Online - Curso Endocrino - Farmacología y Suplementación DeportivaBrianNoch keine Bewertungen
- CuadroDokument7 SeitenCuadroCabrera Sandiego AlexiaNoch keine Bewertungen
- Células endoteliales: estructura, receptores inmunológicos y funcionesDokument3 SeitenCélulas endoteliales: estructura, receptores inmunológicos y funcionesDavid YepezNoch keine Bewertungen
- Dieta AbcdeDokument15 SeitenDieta AbcdeJuan PawneesNoch keine Bewertungen
- Aplicaciones - Microarrays1Dokument0 SeitenAplicaciones - Microarrays1Victor Vasquez MatsudaNoch keine Bewertungen
- Contracción de Fibras Musculares EsqueléticasDokument13 SeitenContracción de Fibras Musculares EsqueléticasAriadna Hildegarda Ayuso ReyesNoch keine Bewertungen
- Mi Parte de InmundoDokument6 SeitenMi Parte de InmundoÓckap OsmaNoch keine Bewertungen
- Grupos Sanguineos 1Dokument2 SeitenGrupos Sanguineos 1Juliana SalcedoNoch keine Bewertungen
- 10 Analgesicos OpioidesDokument10 Seiten10 Analgesicos OpioidesBrayan BonillaNoch keine Bewertungen
- 1° Parcial Bioquímica (Guía-1)Dokument11 Seiten1° Parcial Bioquímica (Guía-1)Isaac HerreraNoch keine Bewertungen