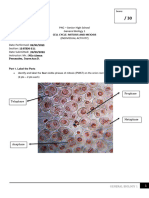
Das könnte Ihnen auch gefallen
- MegaDokument10 SeitenMegaolivia pangaribuanNoch keine Bewertungen
- MSM 217Dokument15 SeitenMSM 217Kaixian LiuNoch keine Bewertungen
- Mitotic Chromosome Doubling of Plant Tissues in VitroDokument15 SeitenMitotic Chromosome Doubling of Plant Tissues in VitroTharani palanisamyNoch keine Bewertungen
- Spermatogonial Stem Cells: Questions, Models and PerspectivesDokument8 SeitenSpermatogonial Stem Cells: Questions, Models and PerspectivesKsenia KamenskaiaNoch keine Bewertungen
- Mitosis & Meiosis NotesDokument6 SeitenMitosis & Meiosis NotesChris_Barber09100% (1)
- MeiosisDokument2 SeitenMeiosisAishatu Adamu GadoNoch keine Bewertungen
- TD1 - Article mAC1Dokument14 SeitenTD1 - Article mAC1sextansNoch keine Bewertungen
- (Biology Form 4) MeiosisDokument4 Seiten(Biology Form 4) MeiosisUmi Zaheera AzharNoch keine Bewertungen
- Meiosis: Week 4 DiscussionDokument9 SeitenMeiosis: Week 4 DiscussionJacob CarlNoch keine Bewertungen
- Chapter 5 Cell Division: Chapt 5:1/5Dokument5 SeitenChapter 5 Cell Division: Chapt 5:1/5ctazimaNoch keine Bewertungen
- 1.3 Cell Division, Diversity and OrganisationDokument12 Seiten1.3 Cell Division, Diversity and OrganisationMo_Bash1Noch keine Bewertungen
- DNA Methylation and Epigenetic Inheritance During Plant GametogenesisDokument9 SeitenDNA Methylation and Epigenetic Inheritance During Plant GametogenesisLili CabNoch keine Bewertungen
- Vic Sample Notes BiologyDokument20 SeitenVic Sample Notes BiologyEvangelene Esquillo SanaNoch keine Bewertungen
- Erq 191Dokument13 SeitenErq 191Nur JannahNoch keine Bewertungen
- Meiosis (: ListenDokument13 SeitenMeiosis (: ListenmuhammadismailNoch keine Bewertungen
- Cytological Bases of HeredityDokument42 SeitenCytological Bases of Heredity'tephanie Salcedo100% (1)
- Biology (Form 4)Dokument25 SeitenBiology (Form 4)Aloysius RajNoch keine Bewertungen
- Biology Module 2010 SeratasDokument28 SeitenBiology Module 2010 SeratasNurul AzuwinNoch keine Bewertungen
- Meiosis - WikipediaDokument31 SeitenMeiosis - WikipediaMustapha AbdullateefNoch keine Bewertungen
- Week 5 Mitosis and MiosisDokument11 SeitenWeek 5 Mitosis and MiosisEarl RénzNoch keine Bewertungen
- PolyploidyDokument7 SeitenPolyploidyleticiabilboNoch keine Bewertungen
- Bioinfo Lect 5Dokument53 SeitenBioinfo Lect 5Khyal DaveNoch keine Bewertungen
- Plant Physiol. 2007 Sprent 575 81Dokument7 SeitenPlant Physiol. 2007 Sprent 575 81Eduardo Antonio Molinari NovoaNoch keine Bewertungen
- Cell Bio ReportDokument5 SeitenCell Bio ReportjonaschewNoch keine Bewertungen
- IntroductionDokument7 SeitenIntroductionNur Hidayatul Aliaa JustinNoch keine Bewertungen
- Genetics Lab 1-7Dokument8 SeitenGenetics Lab 1-7Beatrice Del Rosario100% (2)
- Exam 1 - Essential QuestionsDokument6 SeitenExam 1 - Essential QuestionsMartin De Haro GarciaNoch keine Bewertungen
- International Journal For ParasitologyDokument10 SeitenInternational Journal For ParasitologyGabriel AvilaNoch keine Bewertungen
- TB20003 Lab Manual GeneticsDokument18 SeitenTB20003 Lab Manual Geneticsluver_lynz903814Noch keine Bewertungen
- Mitosis and Meiosis Pre-WorksheetDokument5 SeitenMitosis and Meiosis Pre-WorksheetRoshae SinclairNoch keine Bewertungen
- Symmetry, Asymmetry, and The Cell Cycle in Plants: Known Knowns and Some Known UnknownsDokument11 SeitenSymmetry, Asymmetry, and The Cell Cycle in Plants: Known Knowns and Some Known Unknownsshiksha torooNoch keine Bewertungen
- Mitosis Chromosomes and Chromosomal Number: Form 4 Biology Chapter 5 Cell DivisionDokument27 SeitenMitosis Chromosomes and Chromosomal Number: Form 4 Biology Chapter 5 Cell DivisionMei QiiNoch keine Bewertungen
- Meiosis & MitosisDokument8 SeitenMeiosis & MitosisAna Melisa DorueloNoch keine Bewertungen
- Sexual ReproductionDokument4 SeitenSexual Reproductionmelvinkpaamos1982Noch keine Bewertungen
- Subhashree Parida 2101020204-AssignmentDokument8 SeitenSubhashree Parida 2101020204-AssignmentHero PratikNoch keine Bewertungen
- To Be or Not To Be The Odd One Out - Allele-Specific Transcription in Pentaploid Dogroses (Rosa L. Sect. Caninae (DC.) Ser)Dokument15 SeitenTo Be or Not To Be The Odd One Out - Allele-Specific Transcription in Pentaploid Dogroses (Rosa L. Sect. Caninae (DC.) Ser)Ahmad LabibNoch keine Bewertungen
- Marker-Based Analysis of Genome Structure and DNA Methylation in A Watermelon (Citrullus Lanatus) Ploidy Series Chun Guo WANG Et Al. 2009. RCDokument14 SeitenMarker-Based Analysis of Genome Structure and DNA Methylation in A Watermelon (Citrullus Lanatus) Ploidy Series Chun Guo WANG Et Al. 2009. RCelirayenNoch keine Bewertungen
- Bio 215 Notes Meiosis and MitosisDokument17 SeitenBio 215 Notes Meiosis and MitosisPrecious ObhasedoNoch keine Bewertungen
- Esquema de Puncion de Nematodos, EpidermisDokument11 SeitenEsquema de Puncion de Nematodos, EpidermissebayferNoch keine Bewertungen
- Science Half Yearly NotesDokument14 SeitenScience Half Yearly NotesanonNoch keine Bewertungen
- Solution Manual For Essentials of Biology 5th EditionDokument8 SeitenSolution Manual For Essentials of Biology 5th EditionDeanBucktdjx100% (33)
- AP Biology Lab 3 QuestionsDokument3 SeitenAP Biology Lab 3 Questionsedwarddame100% (1)
- Continuity and Variation: Section C of The CSEC Biology SyllabusDokument40 SeitenContinuity and Variation: Section C of The CSEC Biology SyllabusYanique JonesNoch keine Bewertungen
- Cold Spring Harb Perspect Biol-2015-Ohkura-a015859Dokument15 SeitenCold Spring Harb Perspect Biol-2015-Ohkura-a015859wafiqahNoch keine Bewertungen
- Bangs 2000Dokument12 SeitenBangs 2000laciyeg352Noch keine Bewertungen
- Gen. Biology Module 4 Week 7 & 8Dokument3 SeitenGen. Biology Module 4 Week 7 & 8LouisseNoch keine Bewertungen
- Biological Science Let2017Dokument179 SeitenBiological Science Let2017Janine Cudal QuizmundoNoch keine Bewertungen
- Cell Division and Its Regulation - Part 1Dokument80 SeitenCell Division and Its Regulation - Part 1Arslan AmeenNoch keine Bewertungen
- Mitosis Vs MeiosisDokument7 SeitenMitosis Vs MeiosisJD DX100% (1)
- Translation: Perpetuation of Oppression Occurs When One Dictator After Another Seizes Control of ADokument4 SeitenTranslation: Perpetuation of Oppression Occurs When One Dictator After Another Seizes Control of AAnonymous xGIQknNoch keine Bewertungen
- Dolly OkDokument29 SeitenDolly OkDaniel DerianNoch keine Bewertungen
- Bio 267 First SemDokument8 SeitenBio 267 First SemVreen Pietro Mari EstimoNoch keine Bewertungen
- Mitosis and MeiosisDokument8 SeitenMitosis and Meiosisabisantiago6131Noch keine Bewertungen
- Heredity and Variation: 1. Cell DivisionDokument25 SeitenHeredity and Variation: 1. Cell DivisionRosalmi AyuNoch keine Bewertungen
- Genetics of Dioecy and Causal Sex Chromosomes in PlantsDokument37 SeitenGenetics of Dioecy and Causal Sex Chromosomes in PlantsFranco SantinNoch keine Bewertungen
- Module 1: Introduction To GeneticsDokument3 SeitenModule 1: Introduction To Geneticsrizza reyesNoch keine Bewertungen
- Drill 2 CELL CYCLEDokument4 SeitenDrill 2 CELL CYCLEJoyce FernandezNoch keine Bewertungen
- Topical Guidebook For GCE O Level Biology 3 Part 2Von EverandTopical Guidebook For GCE O Level Biology 3 Part 2Bewertung: 5 von 5 Sternen5/5 (1)
- James 1980 Chap44 SugarcaneDokument13 SeitenJames 1980 Chap44 SugarcaneAJMRNoch keine Bewertungen
- UnderwoodDokument2 SeitenUnderwoodAJMRNoch keine Bewertungen
- Allan 1980 Chap51 WheatDokument12 SeitenAllan 1980 Chap51 WheatAJMRNoch keine Bewertungen
- Auckland 1980 ASA Chap15 ChickpeaDokument11 SeitenAuckland 1980 ASA Chap15 ChickpeaAJMRNoch keine Bewertungen
- Alfred 1980 Chap27 HopDokument14 SeitenAlfred 1980 Chap27 HopAJMRNoch keine Bewertungen
- Barnes 1980 ASA Chap09 AlfalfaDokument11 SeitenBarnes 1980 ASA Chap09 AlfalfaAJMRNoch keine Bewertungen
- Maguire 1988 Gena Near-Normal Chiasma Formation and Maintenance in A Short Distal Translocated Segment in MaizeDokument4 SeitenMaguire 1988 Gena Near-Normal Chiasma Formation and Maintenance in A Short Distal Translocated Segment in MaizeAJMRNoch keine Bewertungen
- Badouin 2017 Nat The Sunflower Geno Prov inDokument14 SeitenBadouin 2017 Nat The Sunflower Geno Prov inAJMRNoch keine Bewertungen
- UnderwoodDokument2 SeitenUnderwoodAJMRNoch keine Bewertungen
- Caetano-Pereira 1999 GMB Cell Fusion and Chromatin Degeneration in An Inbred Line of MaizeDokument7 SeitenCaetano-Pereira 1999 GMB Cell Fusion and Chromatin Degeneration in An Inbred Line of MaizeAJMRNoch keine Bewertungen
- Rai 2010 TsitolG Induced Cytomictic Diversity in ZM InbredDokument5 SeitenRai 2010 TsitolG Induced Cytomictic Diversity in ZM InbredAJMRNoch keine Bewertungen
- Allopolyploid Origin of Cardamine Silana (Brassicaceae) From Calabria (Southern Italy) - Karyological, Morphological and Molecular EvidenceDokument16 SeitenAllopolyploid Origin of Cardamine Silana (Brassicaceae) From Calabria (Southern Italy) - Karyological, Morphological and Molecular EvidenceAJMRNoch keine Bewertungen
- Staton 2012 PLJ The Ha Genome Reflects A Recent History of Biased Accum of TEDokument12 SeitenStaton 2012 PLJ The Ha Genome Reflects A Recent History of Biased Accum of TEAJMRNoch keine Bewertungen
- Weber 1981 EHP Maize Pollen Test Systems To Detect NondisjunctionDokument7 SeitenWeber 1981 EHP Maize Pollen Test Systems To Detect NondisjunctionAJMRNoch keine Bewertungen
- Dale 1974 Wats Var Within C Pratensis in EnglandDokument17 SeitenDale 1974 Wats Var Within C Pratensis in EnglandAJMRNoch keine Bewertungen
- Yu 2000 JCB Functional Redundancy in The Maize Meiotic KinetochoreDokument11 SeitenYu 2000 JCB Functional Redundancy in The Maize Meiotic KinetochoreAJMRNoch keine Bewertungen
- Maguire 1962 SC Pachytene and Diakinesis Behavior of The Isochrom 6 of ZMDokument3 SeitenMaguire 1962 SC Pachytene and Diakinesis Behavior of The Isochrom 6 of ZMAJMRNoch keine Bewertungen
- Khatri 1987 Cardamine Pratensis Complex in The Asian Territory of The USSRDokument7 SeitenKhatri 1987 Cardamine Pratensis Complex in The Asian Territory of The USSRAJMRNoch keine Bewertungen
- Lawrence 1931 Genetica The Chromosome Constitution of Cardamine Pratensis and Verbascum PhoeniceumDokument26 SeitenLawrence 1931 Genetica The Chromosome Constitution of Cardamine Pratensis and Verbascum PhoeniceumAJMRNoch keine Bewertungen
- Daniel Todes-Ivan Pavlov - Exploring The Animal Machine (Oxford Portraits in Science) - Oxford University Press, USA (2000)Dokument113 SeitenDaniel Todes-Ivan Pavlov - Exploring The Animal Machine (Oxford Portraits in Science) - Oxford University Press, USA (2000)Miguel NevesNoch keine Bewertungen
- Zhou 2013 SC Molecular Basis of Age-Dependent Vernalization in Cardamine Flexuosa - SMDokument32 SeitenZhou 2013 SC Molecular Basis of Age-Dependent Vernalization in Cardamine Flexuosa - SMAJMRNoch keine Bewertungen
- Zhou 2013 SC Molecular Basis of Age-Dependent Vernalization in Cardamine FlexuosaDokument5 SeitenZhou 2013 SC Molecular Basis of Age-Dependent Vernalization in Cardamine FlexuosaAJMRNoch keine Bewertungen
- Biogeography and Phylogeny of Cardamine (Brassicaceae)Dokument23 SeitenBiogeography and Phylogeny of Cardamine (Brassicaceae)AJMRNoch keine Bewertungen
- Chan 1998 JCS Maize Meiotic Spindles Assemble Around ChromDokument9 SeitenChan 1998 JCS Maize Meiotic Spindles Assemble Around ChromAJMRNoch keine Bewertungen
- Howard 1951 Nat The So-Called Chromosome-Races of Cardamine Pratensis and Nasturtium OfficinaleDokument2 SeitenHoward 1951 Nat The So-Called Chromosome-Races of Cardamine Pratensis and Nasturtium OfficinaleAJMRNoch keine Bewertungen
- Rebernig 2015 PLoSG Non-Recip Intersps Hybridiz Barriers in The Capsella Genus Are Established in The EndospermDokument19 SeitenRebernig 2015 PLoSG Non-Recip Intersps Hybridiz Barriers in The Capsella Genus Are Established in The EndospermAJMRNoch keine Bewertungen
- Sicard 2015 NatCom Divergent Sorting of A Balanced Ancestral Polymorphism Underlies The Establishment of Gene-Flow Barriers in CapsellaDokument10 SeitenSicard 2015 NatCom Divergent Sorting of A Balanced Ancestral Polymorphism Underlies The Establishment of Gene-Flow Barriers in CapsellaAJMRNoch keine Bewertungen
- Souges 3Dokument1 SeiteSouges 3AJMRNoch keine Bewertungen
- Slotte 2013 NatG The Capsella Rubella Genome and The Genomic Conseq of Rapid Mating System Evol-S1Dokument53 SeitenSlotte 2013 NatG The Capsella Rubella Genome and The Genomic Conseq of Rapid Mating System Evol-S1AJMRNoch keine Bewertungen
- Slotte 2013 NatG The Capsella Rubella Genome and The Genomic Conseq of Rapid Mating System EvolDokument7 SeitenSlotte 2013 NatG The Capsella Rubella Genome and The Genomic Conseq of Rapid Mating System EvolAJMRNoch keine Bewertungen
- Biohermes Sars-Cov-2 Neutralizing Antibodies Test Kit Clinical PerformanceDokument2 SeitenBiohermes Sars-Cov-2 Neutralizing Antibodies Test Kit Clinical PerformanceanggialwieNoch keine Bewertungen
- Tools in BioinformaticsDokument17 SeitenTools in BioinformaticsJane Guiron AballaNoch keine Bewertungen
- Bio 152 Adjusted Syllabus Principles of Molecular Biology and BiotechnologyDokument3 SeitenBio 152 Adjusted Syllabus Principles of Molecular Biology and BiotechnologysofeeNoch keine Bewertungen
- Classification Notes PDFDokument20 SeitenClassification Notes PDFBobNoch keine Bewertungen
- 2018 - Circulating microRNAs As Novel Biomarkers of Alzheimer's DiseaseDokument6 Seiten2018 - Circulating microRNAs As Novel Biomarkers of Alzheimer's DiseaseM JNoch keine Bewertungen
- ABO Blood Type WorksheetDokument2 SeitenABO Blood Type WorksheetKrista Vicente88% (8)
- Report in Gen BioDokument7 SeitenReport in Gen BioJavier KennnyNoch keine Bewertungen
- Biomarkers Diagnostic ProgramDokument23 SeitenBiomarkers Diagnostic ProgramAndraReiNoch keine Bewertungen
- Dna Replication: By: Group 3Dokument26 SeitenDna Replication: By: Group 3Jesse Kate GonzalesNoch keine Bewertungen
- Biochemistry Notes - Nucleotide MetabolismDokument5 SeitenBiochemistry Notes - Nucleotide MetabolismneedthisforepiNoch keine Bewertungen
- Doctors of DelhiDokument137 SeitenDoctors of DelhiAkash Panwar83% (6)
- Regeneration. The Processes of Adaptation and Compensation: Phd. Oksana V. KaluzhynaDokument70 SeitenRegeneration. The Processes of Adaptation and Compensation: Phd. Oksana V. Kaluzhynaсветлый поповNoch keine Bewertungen
- Day 5. Warm-Up. STAAR® Blitz. Science. BiologyDokument2 SeitenDay 5. Warm-Up. STAAR® Blitz. Science. BiologyRaul Ramirez RangelNoch keine Bewertungen
- Cell Biology and GeneticsDokument54 SeitenCell Biology and Geneticspnguyen810Noch keine Bewertungen
- PharmacokintaticDokument38 SeitenPharmacokintaticnoor norNoch keine Bewertungen
- Zydus Cadila: Company Overview and Regulatory PositionDokument26 SeitenZydus Cadila: Company Overview and Regulatory PositionRupam KunduNoch keine Bewertungen
- Rev Choi2019 Micromachines 11 00021 v2Dokument20 SeitenRev Choi2019 Micromachines 11 00021 v2Alejandra CastillaNoch keine Bewertungen
- Learn Vibrant: Biotechnology Principles and Processes Class 12 Biology Revision NotesDokument22 SeitenLearn Vibrant: Biotechnology Principles and Processes Class 12 Biology Revision NotesAmaan KhanNoch keine Bewertungen
- Certificate: Bagus Ramanto SaputraDokument2 SeitenCertificate: Bagus Ramanto SaputraBagus Ramanto SaputraNoch keine Bewertungen
- Nutrigenomics The Genome Food InterfaceDokument8 SeitenNutrigenomics The Genome Food InterfaceElisua G.I.Noch keine Bewertungen
- Plant Cell CultureDokument8 SeitenPlant Cell Culturenurul9535Noch keine Bewertungen
- Describe Five Examples of How Each Level of Disease Prevention Is Practiced in Your CountryDokument1 SeiteDescribe Five Examples of How Each Level of Disease Prevention Is Practiced in Your Countryhasfa konsoNoch keine Bewertungen
- Genetically Modified FoodsDokument6 SeitenGenetically Modified FoodsBicycle ThiefNoch keine Bewertungen
- Amoeba Sisters Video Recap: Dichotomous Keys With Scientific NamesDokument2 SeitenAmoeba Sisters Video Recap: Dichotomous Keys With Scientific NamesDaniel Pinzon56% (9)
- BT I 2018 Revision Question Bank Class Xi: Living World and Biological ClassificationDokument8 SeitenBT I 2018 Revision Question Bank Class Xi: Living World and Biological ClassificationNaman BobalNoch keine Bewertungen
- Mardinoglu 2014Dokument14 SeitenMardinoglu 2014Julia SCNoch keine Bewertungen
- AEM3540 Management Score Sheet and Examiner BreifDokument5 SeitenAEM3540 Management Score Sheet and Examiner BreifAbdullah Al MasumNoch keine Bewertungen
- CHAPTER Acellular LifeDokument4 SeitenCHAPTER Acellular LifeRahila AkramNoch keine Bewertungen
- Fsba Module 2 Unit 1 Notes EnglishDokument19 SeitenFsba Module 2 Unit 1 Notes EnglishDina HushamNoch keine Bewertungen
- 04 Physiology of The Pancreas PDFDokument66 Seiten04 Physiology of The Pancreas PDFMonesa Christy VillanuevaNoch keine Bewertungen