Das könnte Ihnen auch gefallen
- TEMA 2.2 Fisiología Del Sistema Esquelético PDFDokument75 SeitenTEMA 2.2 Fisiología Del Sistema Esquelético PDFcgilgasNoch keine Bewertungen
- BIO2BAC Cuestiones Temas 1 5Dokument24 SeitenBIO2BAC Cuestiones Temas 1 5cgilgasNoch keine Bewertungen
- Cuestiones Tema 3Dokument1 SeiteCuestiones Tema 3cgilgasNoch keine Bewertungen
- Tema 10 - Los ConductismosDokument16 SeitenTema 10 - Los ConductismosUnai NoizeNoch keine Bewertungen
- Brisa MarinaDokument22 SeitenBrisa MarinaDaniel BreraNoch keine Bewertungen
- Recetas para Prepar Un Menu PDFDokument36 SeitenRecetas para Prepar Un Menu PDFcgilgasNoch keine Bewertungen
- Ud1-Glosario BiologíaDokument3 SeitenUd1-Glosario BiologíacgilgasNoch keine Bewertungen
- Coronavirus FPBDokument1 SeiteCoronavirus FPBcgilgasNoch keine Bewertungen
- CINE Y MATES EjerciciosDokument9 SeitenCINE Y MATES EjercicioscgilgasNoch keine Bewertungen
- Resumen Geomorofologia PDFDokument365 SeitenResumen Geomorofologia PDFcgilgasNoch keine Bewertungen
- CGG Tema 22Dokument12 SeitenCGG Tema 22cgilgasNoch keine Bewertungen
- Issue36 Infect Rolecards SpanishDokument2 SeitenIssue36 Infect Rolecards SpanishcgilgasNoch keine Bewertungen
- David-Origen Universo y TecnologiaDokument2 SeitenDavid-Origen Universo y TecnologiacgilgasNoch keine Bewertungen
- Decreto 108-Valencia Curriculo de PrimariaDokument370 SeitenDecreto 108-Valencia Curriculo de PrimariaAitana2013Noch keine Bewertungen
- 2ºeso-UD1-4 REPASODokument4 Seiten2ºeso-UD1-4 REPASOcgilgasNoch keine Bewertungen
- Problemas Aa Resueltos PDFDokument5 SeitenProblemas Aa Resueltos PDFcgilgasNoch keine Bewertungen
- Anatomía Aplic PDFDokument10 SeitenAnatomía Aplic PDFcgilgasNoch keine Bewertungen
- CGG Tema 12Dokument8 SeitenCGG Tema 12cgilgasNoch keine Bewertungen
- BriofitosDokument56 SeitenBriofitosMiguel BueuNoch keine Bewertungen
- TEMA 3 - Glucidos y Lípidos - Mapa MentalDokument4 SeitenTEMA 3 - Glucidos y Lípidos - Mapa MentalcgilgasNoch keine Bewertungen
- NeurohistogénesisDokument20 SeitenNeurohistogénesiscgilgas100% (1)
- Tema 16-Bio y GeoDokument20 SeitenTema 16-Bio y GeocgilgasNoch keine Bewertungen
- ConnectorsDokument5 SeitenConnectorscgilgasNoch keine Bewertungen
- Examen Final Citologia e HistologiaDokument4 SeitenExamen Final Citologia e HistologiacgilgasNoch keine Bewertungen
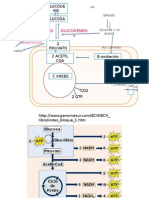
- MetabolismoDokument30 SeitenMetabolismocgilgasNoch keine Bewertungen
- Tema EvolucionDokument34 SeitenTema EvolucionJosema GjNoch keine Bewertungen
- Cuestionario MicrobiologiaDokument10 SeitenCuestionario MicrobiologiacgilgasNoch keine Bewertungen
- Ejercicios Turbidimetría y NefelometríaDokument2 SeitenEjercicios Turbidimetría y Nefelometríachorvo100% (1)
- Biosferas y BiomasDokument7 SeitenBiosferas y BiomasLeyda EscorcheNoch keine Bewertungen
- Cuarto Apuntes Sem 7Dokument81 SeitenCuarto Apuntes Sem 7SANTIAGO ANDRES PLAZAS MARTINEZNoch keine Bewertungen
- Adaptacion Al EntrenamientoDokument20 SeitenAdaptacion Al EntrenamientoCami CamiNoch keine Bewertungen
- Avances Tecnológicos en El Tratamiento de Enfermedades RespiratoriasDokument3 SeitenAvances Tecnológicos en El Tratamiento de Enfermedades RespiratoriasLuz Mery RGNoch keine Bewertungen
- Herencia TransgresivaDokument7 SeitenHerencia TransgresivaWillSmith SanchezNoch keine Bewertungen
- Iip Actividad #1 Biologia-9 ListoDokument4 SeitenIip Actividad #1 Biologia-9 ListoAuri VasquezNoch keine Bewertungen
- CRONONUTRICIÓNDokument4 SeitenCRONONUTRICIÓNMariaNoch keine Bewertungen
- 14 Gubiologia 7Dokument2 Seiten14 Gubiologia 7JonnieAriasFNoch keine Bewertungen
- Problemas de Diluciones AvanzadoDokument3 SeitenProblemas de Diluciones Avanzadoargentina-2009Noch keine Bewertungen
- Guia 3 Bioquímica Grado 10° Del 1 Al 27 de JunioDokument6 SeitenGuia 3 Bioquímica Grado 10° Del 1 Al 27 de JunioBiologiaFaustinistaNoch keine Bewertungen
- Glosario para Entender La Sociología Figuracional PDFDokument41 SeitenGlosario para Entender La Sociología Figuracional PDFCarolina ChaparroNoch keine Bewertungen
- Introducción A La BioquímicaDokument31 SeitenIntroducción A La BioquímicaEnos DEGBENoch keine Bewertungen
- Triptico Sistema Nervioso CentralDokument3 SeitenTriptico Sistema Nervioso CentralNuri AmrNoch keine Bewertungen
- Modelo de TripticoDokument2 SeitenModelo de TripticoVictor A. Figueroa Marcos0% (1)
- Organo FlorDokument9 SeitenOrgano FlorsanducanaNoch keine Bewertungen
- Arquitectura de Plantas LeñosasDokument25 SeitenArquitectura de Plantas LeñosaspiperagaufinNoch keine Bewertungen
- Reticularismo o NeuronismoDokument3 SeitenReticularismo o Neuronismofernando lindeNoch keine Bewertungen
- TECNICAS COPROPARASITOSCOPICAS - A-2020 - Guia Complementaria Parasitologia PDFDokument7 SeitenTECNICAS COPROPARASITOSCOPICAS - A-2020 - Guia Complementaria Parasitologia PDFJaver Andres HernandezNoch keine Bewertungen
- GENOMASDokument10 SeitenGENOMASJavi EspinozaNoch keine Bewertungen
- Estrategia - Industrial y Medioambiente Laboral en Ecuador-Raul HarariDokument221 SeitenEstrategia - Industrial y Medioambiente Laboral en Ecuador-Raul HararibermeomNoch keine Bewertungen
- Planeación BGMB-B2 U3Dokument9 SeitenPlaneación BGMB-B2 U3David SanchezNoch keine Bewertungen
- Práctica 2 Esquivel Flores Medina y TaparaDokument18 SeitenPráctica 2 Esquivel Flores Medina y TaparaLuiggi G Quispe RNoch keine Bewertungen
- BIOQUIMICADokument3 SeitenBIOQUIMICAIsaac GómezNoch keine Bewertungen
- Guia Klein de HomosexualidadDokument7 SeitenGuia Klein de HomosexualidadPsCcyVaze100% (1)
- Resumen Reproducción Sexual de Los HongosDokument9 SeitenResumen Reproducción Sexual de Los HongosJuan SandovalNoch keine Bewertungen
- 2 PDFDokument8 Seiten2 PDFNinaquispe Carlita Valeria0% (1)
- Cta2-U1-Sesion 03Dokument10 SeitenCta2-U1-Sesion 03Aaron Chapoñan RomeroNoch keine Bewertungen
- Inmunidad Frente A Virus y ParásitosDokument14 SeitenInmunidad Frente A Virus y ParásitosLiliana JGNoch keine Bewertungen
- Shock SepticoDokument25 SeitenShock SepticoAngelo SarmientoNoch keine Bewertungen