Das könnte Ihnen auch gefallen
- Ejercicio PhysioEx 3 Actividad 6Dokument5 SeitenEjercicio PhysioEx 3 Actividad 6Karla HuamaniNoch keine Bewertungen
- Resultados Preparación de Medios de CultivosDokument8 SeitenResultados Preparación de Medios de CultivosLisandro Alvarez0% (1)
- Acción Enzimatica de La Alfa AmilasaDokument9 SeitenAcción Enzimatica de La Alfa AmilasaFabiola Del Pilar CuroNoch keine Bewertungen
- Practica 5Dokument9 SeitenPractica 5JocelynNoch keine Bewertungen
- Estracción de CaseinaDokument4 SeitenEstracción de Caseinamaria stella ortega suarezNoch keine Bewertungen
- Reporte Cinética Enzimática ACTUAL 2023-1Dokument9 SeitenReporte Cinética Enzimática ACTUAL 2023-1Alta Gracia ArguellesNoch keine Bewertungen
- ISOMERASASDokument9 SeitenISOMERASASAndia Cch100% (1)
- Receptores CelularesDokument36 SeitenReceptores CelularesyovanaNoch keine Bewertungen
- Agua y PHDokument42 SeitenAgua y PHGO NeuroScience GFNoch keine Bewertungen
- Fraccionamiento CelularDokument3 SeitenFraccionamiento CelularreneNoch keine Bewertungen
- PNO Suspension - Equipo 5Dokument12 SeitenPNO Suspension - Equipo 5Monserrat VilaNoch keine Bewertungen
- PRÁCTICA 2 Coprológico y CoproparasitarioDokument8 SeitenPRÁCTICA 2 Coprológico y CoproparasitarioEstefanía Mariño100% (1)
- BioDokument7 SeitenBioViridiana GarcíaNoch keine Bewertungen
- Práctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Dokument11 SeitenPráctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Angel Gustavo HernandezNoch keine Bewertungen
- Precipitación, Separación y Punto Isoeléctrico de Proteínas-Sección ADokument4 SeitenPrecipitación, Separación y Punto Isoeléctrico de Proteínas-Sección ADynarko RuizNoch keine Bewertungen
- La Titulación o Valoración Química Es Un Proceso Por El Que Se Mide La Cantidad o La Concentración de Una Sustancia en Una MuestraDokument1 SeiteLa Titulación o Valoración Química Es Un Proceso Por El Que Se Mide La Cantidad o La Concentración de Una Sustancia en Una MuestraOmaryulys SuberoNoch keine Bewertungen
- ColesterolDokument3 SeitenColesterolPaola Herbas R0% (1)
- Prueba de LowryDokument4 SeitenPrueba de LowryErick Amador GaytánNoch keine Bewertungen
- Inhibicion EnzimaticaDokument14 SeitenInhibicion EnzimaticaFiorella Rosas RuizNoch keine Bewertungen
- Importancia de Las EnzimasDokument6 SeitenImportancia de Las EnzimasYerika Cañizalez de Partidas100% (1)
- Plantilla de PresentaciónDokument35 SeitenPlantilla de PresentaciónLori EsbeltoNoch keine Bewertungen
- 3.permeabilidad de La MembranaDokument20 Seiten3.permeabilidad de La MembranaJunior Rey ColomeNoch keine Bewertungen
- FIBRINOLISISDokument2 SeitenFIBRINOLISISSara JimenezNoch keine Bewertungen
- PRÁCTICA #08 EnzimasDokument6 SeitenPRÁCTICA #08 Enzimasjimmy gomezNoch keine Bewertungen
- Fases de Crecimiento BacterianoDokument3 SeitenFases de Crecimiento BacterianoGiosué Reyes HernandezNoch keine Bewertungen
- IsoenzimasDokument9 SeitenIsoenzimasDavid MercuryNoch keine Bewertungen
- PRACTICA TEST DE EMBARAZO y GLUCOSADokument10 SeitenPRACTICA TEST DE EMBARAZO y GLUCOSArhernandez_884074Noch keine Bewertungen
- Aislamiento de ProteínasDokument2 SeitenAislamiento de ProteínasLeonardo Santizo100% (1)
- Sesion 08. - Guia de Ejercicios N°02Dokument45 SeitenSesion 08. - Guia de Ejercicios N°02riesgos33100% (1)
- 15 Procesamiento Rna - 17 2Dokument32 Seiten15 Procesamiento Rna - 17 2KNayNoch keine Bewertungen
- Modificaciones PostraduccionalesDokument15 SeitenModificaciones PostraduccionalesDaniel Ovando100% (1)
- Mapa PDFDokument1 SeiteMapa PDFedithNoch keine Bewertungen
- Cuestionario Práctica 6 - Técnicas de Tinción DiferencialesDokument4 SeitenCuestionario Práctica 6 - Técnicas de Tinción DiferencialesLizzet SalazarNoch keine Bewertungen
- Laboratorio 3 - Ubicuidad BacterianaDokument7 SeitenLaboratorio 3 - Ubicuidad BacterianaCristina Paez100% (1)
- Informe MonohíbridoDokument6 SeitenInforme Monohíbridosara gonzalezNoch keine Bewertungen
- Discucion Grupal 2019Dokument53 SeitenDiscucion Grupal 2019Ibette NorttonNoch keine Bewertungen
- Tinción de GRAMDokument1 SeiteTinción de GRAMAna Isabel LópezNoch keine Bewertungen
- PhysioEx Ejercicio 5 Actividad 4 (B)Dokument3 SeitenPhysioEx Ejercicio 5 Actividad 4 (B)henryNoch keine Bewertungen
- Prueba de La CatalasaDokument4 SeitenPrueba de La CatalasaJoel H. Perez100% (2)
- Concentración Mínima InhibitoriaDokument55 SeitenConcentración Mínima InhibitoriaNataly Sofia Poma PerezNoch keine Bewertungen
- El Citoesqueleto BacterianoDokument57 SeitenEl Citoesqueleto Bacterianokelly100% (3)
- Control de CC - Biologia PDFDokument8 SeitenControl de CC - Biologia PDFdiscusión en 2 horasNoch keine Bewertungen
- Serie. Termodinámica y CatálisisDokument3 SeitenSerie. Termodinámica y CatálisisEmiliano Ruiz RamirezNoch keine Bewertungen
- Cuadro Comparativo B y CDokument5 SeitenCuadro Comparativo B y CMaria AriasNoch keine Bewertungen
- Enzimas InformeDokument8 SeitenEnzimas InformeFabio BernalNoch keine Bewertungen
- Cuestionario de Antifúngicos y Resistencia A Los Antifúngicos Con RespuestasDokument4 SeitenCuestionario de Antifúngicos y Resistencia A Los Antifúngicos Con RespuestasFrank PinedaNoch keine Bewertungen
- Practica 2-Formas Celulares II-EdgarBZDokument8 SeitenPractica 2-Formas Celulares II-EdgarBZGaby BaltazarNoch keine Bewertungen
- Semana 10 InformeDokument9 SeitenSemana 10 InformejhonatanNoch keine Bewertungen
- Practica #4 Lab BioquimicaDokument5 SeitenPractica #4 Lab BioquimicaLEIDYS CENITH FONSECA TORRESNoch keine Bewertungen
- Universidad Tecnica de AmbatoDokument2 SeitenUniversidad Tecnica de AmbatoAnGiie Moran100% (1)
- Informe de FermentacionDokument24 SeitenInforme de FermentacionSARANoch keine Bewertungen
- Metabolismo Pentobarbital, Ventaja Del Fármaco Con Respecto Al Producto Cabeza de Serie.Dokument4 SeitenMetabolismo Pentobarbital, Ventaja Del Fármaco Con Respecto Al Producto Cabeza de Serie.joget jonesNoch keine Bewertungen
- Factores Que Influyen en El CrecimientoDokument4 SeitenFactores Que Influyen en El Crecimientoangel226293Noch keine Bewertungen
- Acatalasemia IiDokument3 SeitenAcatalasemia IiHugoAnthonyDelgadoCastilloNoch keine Bewertungen
- Informe 5 BiologiaDokument6 SeitenInforme 5 BiologiaYeison DiazNoch keine Bewertungen
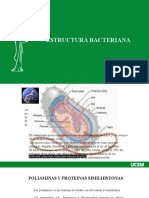
- Estructura Bacteriana Corpusculos Metacromaticos y EsporasDokument22 SeitenEstructura Bacteriana Corpusculos Metacromaticos y EsporasMejia Jhenny YaneNoch keine Bewertungen
- Cinética EnzimaticaDokument23 SeitenCinética EnzimaticaArturo MoraNoch keine Bewertungen
- Marco TeóricoDokument8 SeitenMarco TeóricoDaniel Arroyo AedoNoch keine Bewertungen
- Cinética EnzimáticaDokument6 SeitenCinética EnzimáticaAngie More SolanoNoch keine Bewertungen
- Enzimas Determinación CuantitativaDokument14 SeitenEnzimas Determinación CuantitativaIngeniero PerezNoch keine Bewertungen
- Investigacion Mitosis y MeiosisDokument5 SeitenInvestigacion Mitosis y MeiosisJohan MendezNoch keine Bewertungen
- Taller NO. 1 de Niveles de OrganizacionDokument4 SeitenTaller NO. 1 de Niveles de OrganizacionAna GonzalezNoch keine Bewertungen
- Práctica 12. Digestión y Metabolismo de ProteínasDokument7 SeitenPráctica 12. Digestión y Metabolismo de ProteínasKatherin Arcaya CruzNoch keine Bewertungen
- Metabolismo de Los CarbohidratosDokument33 SeitenMetabolismo de Los CarbohidratosEduardo Perez100% (1)
- Phet Gene-Expression-Essentials-Html-Guide - EsDokument3 SeitenPhet Gene-Expression-Essentials-Html-Guide - EsClauToreNoch keine Bewertungen
- Cloroplastos - Leucoplastos y CromoplastosDokument26 SeitenCloroplastos - Leucoplastos y CromoplastosJorge Jesus Jeri JimenezNoch keine Bewertungen
- Taller Metabolismo de Proteínas 2.0Dokument3 SeitenTaller Metabolismo de Proteínas 2.0Tomás NavasNoch keine Bewertungen
- Genotecas Genomicas de cADNDokument19 SeitenGenotecas Genomicas de cADNLizzete Martinez Ramirez100% (1)
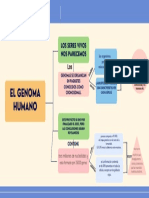
- Genoma HumanoDokument1 SeiteGenoma HumanoKarla Cruzado SilvaNoch keine Bewertungen
- TP Citologia AnimalDokument61 SeitenTP Citologia AnimalGustavo A. SuarezNoch keine Bewertungen
- Choice Sobre Hidratos de CarbonoDokument3 SeitenChoice Sobre Hidratos de CarbonoJack MondragonNoch keine Bewertungen
- Papilas GustativasDokument4 SeitenPapilas GustativaserneyNoch keine Bewertungen
- ENZIMIOLOGÍA - Obstetricia PDFDokument65 SeitenENZIMIOLOGÍA - Obstetricia PDFDerecho IcaNoch keine Bewertungen
- Apunes Virus JuninDokument222 SeitenApunes Virus JuninMerzz DieNoch keine Bewertungen
- Electroforesis de ProteinasDokument1 SeiteElectroforesis de ProteinasCarola Sofía Bahamondes DonosoNoch keine Bewertungen
- Paso 2 Mejoramiento Animal Grupo Colaborativo 201108 9Dokument23 SeitenPaso 2 Mejoramiento Animal Grupo Colaborativo 201108 9Duvan Sneider Lopez GonzalezNoch keine Bewertungen
- Lisosomas, Muerte y Tráfico VesicularDokument66 SeitenLisosomas, Muerte y Tráfico VesicularUSMP FN ARCHIVOS100% (1)
- Radiobiology Book (001-556) (026-031) .En - EsDokument6 SeitenRadiobiology Book (001-556) (026-031) .En - EsOmar Garrido RomeroNoch keine Bewertungen
- 3a Clase ( - 07) - Ac. Nucleicos - Sintesis de Purina y PirimidinaDokument16 Seiten3a Clase ( - 07) - Ac. Nucleicos - Sintesis de Purina y PirimidinaGlenis Acevedo AgredaNoch keine Bewertungen
- Tema 3 CitogeneticaDokument38 SeitenTema 3 CitogeneticaMaría Fernanda Ortiz100% (1)
- Marco TeóricoDokument8 SeitenMarco TeóricoAngela omaña50% (2)
- Juan Toscano Fuenetes, Reparación y Recomendación Del ADNDokument16 SeitenJuan Toscano Fuenetes, Reparación y Recomendación Del ADNJUAN TOSCANONoch keine Bewertungen
- PurinasDokument5 SeitenPurinasGustavo HCNoch keine Bewertungen
- Cuestionarios de TisularDokument13 SeitenCuestionarios de TisularGarcía Pérez Jocelyn ItzelNoch keine Bewertungen
- Laboratorio 3. Extracción de ADN y PCRDokument5 SeitenLaboratorio 3. Extracción de ADN y PCRJuan Esteban Coronado SuescaNoch keine Bewertungen
- Profesora Meri ValdésDokument5 SeitenProfesora Meri ValdéschantelleNoch keine Bewertungen
- Ácidos NucleicosDokument3 SeitenÁcidos NucleicosDaniela Milagros Alvarado NeciosupNoch keine Bewertungen
- LISINADokument1 SeiteLISINAJhonatan IbarraNoch keine Bewertungen
- Replicacion Del Adn en ProcariotasDokument3 SeitenReplicacion Del Adn en ProcariotasPaola Malena Callizaya MariñoNoch keine Bewertungen
- Estructura y Función de La Membrana CelularDokument47 SeitenEstructura y Función de La Membrana CelularlizbethvalenciaNoch keine Bewertungen