Das könnte Ihnen auch gefallen
- Factores AbióticosDokument7 SeitenFactores AbióticosIsaac Herbas MattosNoch keine Bewertungen
- LAB Morfología de Hongos y BacteriasDokument16 SeitenLAB Morfología de Hongos y BacteriasWilliam Reyes PeñaNoch keine Bewertungen
- Camilo Leal Ecosistemas Lénticos y Lóticos 2 PDFDokument5 SeitenCamilo Leal Ecosistemas Lénticos y Lóticos 2 PDFCamilo Leal100% (1)
- 001 Fisica Informe de Caja de ResonanciaDokument6 Seiten001 Fisica Informe de Caja de ResonanciaYolimarVelezNoch keine Bewertungen
- Tubo de Kundt PDFDokument12 SeitenTubo de Kundt PDFMarisol Ortiz GNoch keine Bewertungen
- Responsabilidad Del Educador SujetoDokument4 SeitenResponsabilidad Del Educador SujetoPamela GarciaNoch keine Bewertungen
- Planteamiento Del ProblemaDokument6 SeitenPlanteamiento Del ProblemaLuis OdicioNoch keine Bewertungen
- Maloka PDFDokument15 SeitenMaloka PDFjavier bNoch keine Bewertungen
- Trabajo Final InvestigativoDokument44 SeitenTrabajo Final Investigativoorleidys gomez caicedoNoch keine Bewertungen
- Maloka ElectromagnetismoDokument10 SeitenMaloka ElectromagnetismoSergio Daniel Rodriguez Preciado0% (1)
- Trabajo Practico Péndulo SimpleDokument4 SeitenTrabajo Practico Péndulo SimpleLeonardo A. FunesNoch keine Bewertungen
- Una Visión Holistica de La Investigación PedagógicaDokument4 SeitenUna Visión Holistica de La Investigación PedagógicaMauro MorenoNoch keine Bewertungen
- EJEMPLO-diapositivas Cuasi ExperimentalDokument5 SeitenEJEMPLO-diapositivas Cuasi ExperimentalHebert MamaniNoch keine Bewertungen
- Programa NanomaterialesDokument8 SeitenPrograma NanomaterialesMaxi GonzalezNoch keine Bewertungen
- Taller Moléculas y Seres Vivos Juana FDokument5 SeitenTaller Moléculas y Seres Vivos Juana FJUANA VALENTINA FRANCO CORREDORNoch keine Bewertungen
- Introducción de La MitosisDokument5 SeitenIntroducción de La MitosisSneyder Leyton Cárdenas100% (1)
- Philippe Meirieu Transformacion PedagogicaDokument12 SeitenPhilippe Meirieu Transformacion Pedagogicajairo alberto galindo cuestaNoch keine Bewertungen
- Informe Del OjoDokument4 SeitenInforme Del OjoJonathan Enríquez0% (1)
- ProtoplasmaDokument9 SeitenProtoplasmaChristopher Guevara CheNoch keine Bewertungen
- Propiedades de Las Células CancerigenasDokument9 SeitenPropiedades de Las Células CancerigenasfausNoch keine Bewertungen
- Indices de Diversidad EcosistemicaDokument2 SeitenIndices de Diversidad EcosistemicaJuan Carlos Giraldo RiosNoch keine Bewertungen
- Practica 1 Fuerza ElectricaDokument11 SeitenPractica 1 Fuerza ElectricaCessar Augusto Vergara ArdilaNoch keine Bewertungen
- Ondas Estacionarias en Tubos AbiertosDokument7 SeitenOndas Estacionarias en Tubos AbiertosFrancy Cristina Ramirez BedoyaNoch keine Bewertungen
- Demostracion de Las Leyes de A TermodinamicaDokument20 SeitenDemostracion de Las Leyes de A TermodinamicaJoel Quisuruco CasasNoch keine Bewertungen
- 8a-Campos Magneticos Estaticos - HDokument25 Seiten8a-Campos Magneticos Estaticos - HMario Javier PradaNoch keine Bewertungen
- Manual PlactonDokument60 SeitenManual PlactonNathaliaZarzaNoch keine Bewertungen
- Laboratorio Morfologia Del OjoDokument14 SeitenLaboratorio Morfologia Del OjoNiceth Natalia Ricaurte MorenoNoch keine Bewertungen
- Evolucion de La MitosisDokument5 SeitenEvolucion de La MitosisLina Munoz100% (1)
- Informe Ondas Sonoras PDFDokument17 SeitenInforme Ondas Sonoras PDFJimmy Alexander Ochoa RuizNoch keine Bewertungen
- Fibra OpticaDokument8 SeitenFibra OpticaSergio BoccheciampeNoch keine Bewertungen
- Practica de Laboratorio # 4-Estudio de Las Lentes-2012505Dokument5 SeitenPractica de Laboratorio # 4-Estudio de Las Lentes-2012505Ruben100% (1)
- Impulso NerviosoDokument10 SeitenImpulso NerviosomimiNoch keine Bewertungen
- Estructura Interna de La MateriaDokument4 SeitenEstructura Interna de La MateriaLuis Carlos MosqueraNoch keine Bewertungen
- Guia Basica de Quimica 4Dokument3 SeitenGuia Basica de Quimica 4ovidio martinez100% (1)
- Informe de Extracción de Adn BiologíaDokument5 SeitenInforme de Extracción de Adn BiologíaCarito Lisseth BorjaNoch keine Bewertungen
- Transporte en La MembranaDokument10 SeitenTransporte en La MembranaCRISTIAN MANUEL SAAVEDRA RODRIGUEZNoch keine Bewertungen
- Modelos de ProduccionDokument6 SeitenModelos de Produccionhector cedeñoNoch keine Bewertungen
- Prac 6 Permeabilidad Celular 1Dokument18 SeitenPrac 6 Permeabilidad Celular 1freshaNoch keine Bewertungen
- Tema 3. Señalización Celular PDFDokument18 SeitenTema 3. Señalización Celular PDFAelita RuseelNoch keine Bewertungen
- Microscopia de Contraste de FasesDokument7 SeitenMicroscopia de Contraste de FasesIvan Salas Gaeta100% (1)
- Practica OsmosisDokument6 SeitenPractica OsmosisJhonatan Santos MartínezNoch keine Bewertungen
- Fenómenos de MembranasDokument8 SeitenFenómenos de MembranasArlex Lara Sánchez100% (1)
- Articulo Cientifico Reconocimiento y Clasificacion de SuelosDokument14 SeitenArticulo Cientifico Reconocimiento y Clasificacion de SuelosAnyelii08Noch keine Bewertungen
- Secuencia 2° Digestivo Uruguay EducaDokument9 SeitenSecuencia 2° Digestivo Uruguay EducaIanara RodríguezNoch keine Bewertungen
- La MitocondriaDokument4 SeitenLa MitocondriaDavid GatoNoch keine Bewertungen
- Laboratorio06 Osmosis en Plantas y AnimalesDokument6 SeitenLaboratorio06 Osmosis en Plantas y AnimalesSalomón Marín GarnicaNoch keine Bewertungen
- Informe MicrobiologíaDokument5 SeitenInforme MicrobiologíaLiizhiiz Vaca100% (1)
- Sistema OsteoartromuscularDokument5 SeitenSistema OsteoartromuscularAdriánGonzálezNoch keine Bewertungen
- Morbilidad Infantil PDFDokument27 SeitenMorbilidad Infantil PDFRebeca ChacínNoch keine Bewertungen
- Teorías Sobre El Origen de La Célula EucariotaDokument1 SeiteTeorías Sobre El Origen de La Célula EucariotaMauro Villarrubia67% (3)
- Encéfalo: Prosencéfalo, Mesencéfalo, RombencéfaloDokument13 SeitenEncéfalo: Prosencéfalo, Mesencéfalo, RombencéfaloSthefanny MedinaNoch keine Bewertungen
- Informe de Laboratorio de Biología - MicroscopíaDokument11 SeitenInforme de Laboratorio de Biología - MicroscopíameromachofullNoch keine Bewertungen
- El Sistema o Aparato Excretor de Los AnimalesDokument5 SeitenEl Sistema o Aparato Excretor de Los AnimalesEvelynNoch keine Bewertungen
- Los Cinco ReinosDokument12 SeitenLos Cinco ReinosJhoselyn Esther GarcíaNoch keine Bewertungen
- Anelidos Informe Nota 95Dokument8 SeitenAnelidos Informe Nota 95Noemi DuranNoch keine Bewertungen
- Quiralidad en La Química La Física y La Biología: by Jean Paul Tunjano CallejasDokument8 SeitenQuiralidad en La Química La Física y La Biología: by Jean Paul Tunjano CallejasJean Paul HessNoch keine Bewertungen
- Transporte A Través de La Membrana Celular o PlasmáticaDokument6 SeitenTransporte A Través de La Membrana Celular o PlasmáticaFco ZalgadoNoch keine Bewertungen
- Superficies EquipotencialesDokument10 SeitenSuperficies EquipotencialesGabriel PintoNoch keine Bewertungen
- Celula HipertonicaDokument33 SeitenCelula HipertonicaErika Viviana MoncadaNoch keine Bewertungen
- Contraccion Muscular ExpoDokument20 SeitenContraccion Muscular ExpoElisabet KentNoch keine Bewertungen
- EdulcorantesDokument5 SeitenEdulcorantesviaereaNoch keine Bewertungen
- Seminario Integracion MetabolicaDokument23 SeitenSeminario Integracion MetabolicafawingpeNoch keine Bewertungen
- Fibra DieteticaDokument12 SeitenFibra DieteticaIvette DomínguezNoch keine Bewertungen
- Antropometria Universitarios PDFDokument6 SeitenAntropometria Universitarios PDFRoberto Esparza SolisNoch keine Bewertungen
- Perspectiva y Patrónde AlimentaciDokument5 SeitenPerspectiva y Patrónde AlimentaciRoberto Esparza SolisNoch keine Bewertungen
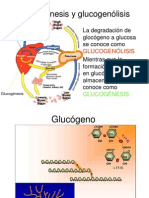
- 11GLUCOGENOLISISDokument30 Seiten11GLUCOGENOLISISRoberto Esparza Solis100% (1)
- Ejercicios de Maximos y MinimosDokument2 SeitenEjercicios de Maximos y MinimosJuan Camilo PardoNoch keine Bewertungen
- Laboratorio 6 FluidosDokument7 SeitenLaboratorio 6 FluidosSnaider Yari SNoch keine Bewertungen
- Herramientas de DibujoDokument6 SeitenHerramientas de DibujoAnthonyCanoNoch keine Bewertungen
- 8.pila ElectroquimicaDokument6 Seiten8.pila ElectroquimicaKELLY JOHANA GOMEZ MEZANoch keine Bewertungen
- Leccion Eva 1Dokument4 SeitenLeccion Eva 1Victor Rodrigo Cruz DelgadoNoch keine Bewertungen
- Graficos Significado EspañolDokument2 SeitenGraficos Significado EspañolGloria Alfonzo100% (2)
- Guía # 4 Funcion Exponencial, Logaritmica Y A TrozosDokument7 SeitenGuía # 4 Funcion Exponencial, Logaritmica Y A TrozosBrayan Stiven Hurtado SamudioNoch keine Bewertungen
- Línea Del TiempoDokument8 SeitenLínea Del TiempoLilián Rodríguez LópezNoch keine Bewertungen
- Comunicación - Eficaz - Escucha ActivaDokument16 SeitenComunicación - Eficaz - Escucha ActivajonashNoch keine Bewertungen
- Práctica #3 (Monitoria) Ejercicios Sin Hacer PDFDokument2 SeitenPráctica #3 (Monitoria) Ejercicios Sin Hacer PDFOsiris Junior TamayoNoch keine Bewertungen
- 13 Trabajo MecánicoDokument9 Seiten13 Trabajo MecánicoDiego FloresNoch keine Bewertungen
- CAPITULO 4 - Radar SecundarioDokument15 SeitenCAPITULO 4 - Radar Secundariogoliat52Noch keine Bewertungen
- Conceptos Básicos de Instrumentación IndustrialDokument10 SeitenConceptos Básicos de Instrumentación IndustrialRoberthGutierrezNoch keine Bewertungen
- Poleas DiapositivasDokument9 SeitenPoleas DiapositivasLuis Fernando Guerra100% (2)
- PH de CañeriaDokument4 SeitenPH de CañeriaHugo OrtegaNoch keine Bewertungen
- Laboratorio Nº7 Asorción de Ácido Oxálico Sobre Carbón Vegetal A Partit de Soluciones AcuosasDokument8 SeitenLaboratorio Nº7 Asorción de Ácido Oxálico Sobre Carbón Vegetal A Partit de Soluciones AcuosasValentina Salazar LópezNoch keine Bewertungen
- Act 8termoDokument10 SeitenAct 8termoalexandra aguirreNoch keine Bewertungen
- Bioelementos Ensayo EmanuelDokument4 SeitenBioelementos Ensayo EmanuelSebastián MendezNoch keine Bewertungen
- Experimento de Conductividad Eléctrica en La Luz EléctricaDokument2 SeitenExperimento de Conductividad Eléctrica en La Luz EléctricaMyu Alove DahabyNoch keine Bewertungen
- Informe N 05 LaborDokument15 SeitenInforme N 05 LaborAlexander SanchezNoch keine Bewertungen
- GLosario de Teoria de SistemasDokument6 SeitenGLosario de Teoria de SistemasNorman Jose FloresNoch keine Bewertungen
- Preguntas Instructor de Vuelo ANACDokument153 SeitenPreguntas Instructor de Vuelo ANACeljonny01Noch keine Bewertungen
- Cuestionario Modelo AtómicoDokument2 SeitenCuestionario Modelo AtómicoNicolas Agustin SavoiaNoch keine Bewertungen
- Protocolo Práctica PotenciometríaDokument4 SeitenProtocolo Práctica PotenciometríaLuis Antonio Pineda RamirezNoch keine Bewertungen
- Enfriamiento de La Leche de VacaDokument27 SeitenEnfriamiento de La Leche de VacaNaly Peres GamarraNoch keine Bewertungen
- Manual Easy Lae17500xbbDokument42 SeitenManual Easy Lae17500xbbJairoLópez67% (3)
- Radiestesia CuanticaDokument155 SeitenRadiestesia CuanticaAdriana Del Rosario Zarate95% (41)
- Práctica #4Dokument13 SeitenPráctica #4Jorge Alberto Moran SilvaNoch keine Bewertungen
- Capitulo 6Dokument16 SeitenCapitulo 6mauro31Noch keine Bewertungen
- Formulario AQDokument5 SeitenFormulario AQDiana CetinaNoch keine Bewertungen