Das könnte Ihnen auch gefallen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Chaudhary Arxiv Vortices DPLDokument10 SeitenChaudhary Arxiv Vortices DPLAdrian ScurtuNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Kaur Dust Vortex DCplasma PoP2015Dokument9 SeitenKaur Dust Vortex DCplasma PoP2015Adrian ScurtuNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Fighting Chess With Hikaru Nakamura - Progress in Chess - Karsten Müller, Raymund Stolze - Ed. Olms - 2012.OCR OPT RDokument234 SeitenFighting Chess With Hikaru Nakamura - Progress in Chess - Karsten Müller, Raymund Stolze - Ed. Olms - 2012.OCR OPT RJosé Del Carmen100% (9)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Coherent Vortices in Strongly Coupled Liquids: Week Ending 1 APRIL 2011Dokument4 SeitenCoherent Vortices in Strongly Coupled Liquids: Week Ending 1 APRIL 2011Adrian ScurtuNoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Muller & Lamprecht - Fundamental Chess EndingsDokument9 SeitenMuller & Lamprecht - Fundamental Chess EndingsAnand RamaduraiNoch keine Bewertungen
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Acsm Review HypertensionDokument21 SeitenAcsm Review HypertensionnamikNoch keine Bewertungen
- Effect of Bhastrika Pranayama and Exercise On Lung.1655Dokument2 SeitenEffect of Bhastrika Pranayama and Exercise On Lung.1655Adrian ScurtuNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Science-Religion Interview With Ravindra PDFDokument7 SeitenScience-Religion Interview With Ravindra PDFAdrian ScurtuNoch keine Bewertungen
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Ticos HighSpeedCameraPlasmas V5febrDokument18 SeitenTicos HighSpeedCameraPlasmas V5febrAdrian ScurtuNoch keine Bewertungen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- 8covlea-1 JOAM 2008 PDFDokument6 Seiten8covlea-1 JOAM 2008 PDFAdrian ScurtuNoch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Tibetan Yoga and Secret DoctrinesDokument441 SeitenTibetan Yoga and Secret Doctrinesmday8386% (7)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Observer in The Quantum ExperimentDokument18 SeitenThe Observer in The Quantum ExperimentAdrian ScurtuNoch keine Bewertungen
- Psychophysiology of Yoga Postures Ancient and Modern Perspectives of AsanasDokument4 SeitenPsychophysiology of Yoga Postures Ancient and Modern Perspectives of AsanasAdrian Scurtu100% (1)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Mental UniverseDokument1 SeiteThe Mental Universemauro.pennacchietti100% (2)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- A Sanskrit Grammar For Beginners 1000213438Dokument330 SeitenA Sanskrit Grammar For Beginners 1000213438psi100% (1)
- Zen Without Zen Masters by Camden Benares Text PDFDokument123 SeitenZen Without Zen Masters by Camden Benares Text PDFAdrian Scurtu100% (1)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Indian Cosmological IdeasDokument12 SeitenIndian Cosmological IdeasHrishikesh BhatNoch keine Bewertungen
- Effect of Bhastrika Pranayama and Exercise On Lung.1655Dokument2 SeitenEffect of Bhastrika Pranayama and Exercise On Lung.1655Adrian ScurtuNoch keine Bewertungen
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Samskrtasubodhini. A Sanskrit Primer. (M.deshpande) (2007)Dokument471 SeitenSamskrtasubodhini. A Sanskrit Primer. (M.deshpande) (2007)sktkoshas96% (46)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Biomechanical Characteristics of The Modified.2680Dokument1 SeiteBiomechanical Characteristics of The Modified.2680Adrian ScurtuNoch keine Bewertungen
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- NPJSCHZ 201547Dokument7 SeitenNPJSCHZ 201547Ignacia Cid PintoNoch keine Bewertungen
- JSSM 15 434 PDFDokument17 SeitenJSSM 15 434 PDFAdrian ScurtuNoch keine Bewertungen
- Brahmachari, Dhirendra - YogaVijnanaDokument84 SeitenBrahmachari, Dhirendra - YogaVijnanalivethelife100% (4)
- Ticos - APL97 - Removal of Floating Dust in Glow Discharge Using Plasma JetDokument3 SeitenTicos - APL97 - Removal of Floating Dust in Glow Discharge Using Plasma JetAdrian ScurtuNoch keine Bewertungen
- (2015) Experimental Demonstration of Martian Soil Simulant Removal From A Surface Using a..RSI (2015)Dokument7 Seiten(2015) Experimental Demonstration of Martian Soil Simulant Removal From A Surface Using a..RSI (2015)Adrian ScurtuNoch keine Bewertungen
- Effect of Bhastrika Pranayama and Exercise On Lung.1655Dokument2 SeitenEffect of Bhastrika Pranayama and Exercise On Lung.1655Adrian ScurtuNoch keine Bewertungen
- Introduction To MATLAB - Sikander M. MirzaDokument45 SeitenIntroduction To MATLAB - Sikander M. MirzaNasir JumaniNoch keine Bewertungen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Zen Without Zen Masters by Camden Benares Text PDFDokument123 SeitenZen Without Zen Masters by Camden Benares Text PDFAdrian Scurtu100% (1)
- (2013) High-Speed Imaging of Dust Particles PDFDokument13 Seiten(2013) High-Speed Imaging of Dust Particles PDFAdrian ScurtuNoch keine Bewertungen
- Guide To ManifestationDokument4 SeitenGuide To ManifestationMONIQUE NICOLE ANTERONoch keine Bewertungen
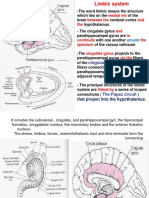
- Limbic SystemDokument18 SeitenLimbic SystemPankajNoch keine Bewertungen
- Mind Does Really Matter: Evidence From Neuroimaging Studies of Emotional Self-Regulation, Psychotherapy, and Placebo EffectDokument19 SeitenMind Does Really Matter: Evidence From Neuroimaging Studies of Emotional Self-Regulation, Psychotherapy, and Placebo Effectseagoddess_simoNoch keine Bewertungen
- 2017 - Vatansever - Default Mode Contributions To Automated Information ProcessingDokument13 Seiten2017 - Vatansever - Default Mode Contributions To Automated Information ProcessingMihaela ChirobociaNoch keine Bewertungen
- Psilocybin S Potential Mechanisms in The Treatment of Depression A Systematic ReviewDokument16 SeitenPsilocybin S Potential Mechanisms in The Treatment of Depression A Systematic ReviewGenis OnnaNoch keine Bewertungen
- Brodmann Areas For HumanDokument32 SeitenBrodmann Areas For HumandyanabellaNoch keine Bewertungen
- Wami Chizarum Project For BuhrecDokument29 SeitenWami Chizarum Project For BuhrecMarvellous OkonobohNoch keine Bewertungen
- Imaging Anatomy Brain and Spine Osborn 1 Ed 2020 PDFDokument3.130 SeitenImaging Anatomy Brain and Spine Osborn 1 Ed 2020 PDFthe gaangster100% (1)
- Northoff - 2022 - Neuropsychoanalysis A Contemporary IntroductionDokument168 SeitenNorthoff - 2022 - Neuropsychoanalysis A Contemporary IntroductionGrzegorz MichalikNoch keine Bewertungen
- Psychology - Theory of Mind - Evolution, Ontogeny, Brain Mechanisms and Psycho PathologyDokument19 SeitenPsychology - Theory of Mind - Evolution, Ontogeny, Brain Mechanisms and Psycho Pathologycdamaia3986Noch keine Bewertungen
- Una Revisión Crítica de Los Criterios de Adicción A Internet Con Sugerencias para El FuturoDokument11 SeitenUna Revisión Crítica de Los Criterios de Adicción A Internet Con Sugerencias para El FuturoANDREA MIKAELA GONZALEZ VERANoch keine Bewertungen
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- 10 1111@j 1460-9568 2006 04785 X PDFDokument13 Seiten10 1111@j 1460-9568 2006 04785 X PDFTlaloc GonzalezNoch keine Bewertungen
- The Pathophysiology of Schizophrenia DisordersDokument16 SeitenThe Pathophysiology of Schizophrenia DisordersElsya ApriliaNoch keine Bewertungen
- Feature Article The Neurobiology of Thought: The Groundbreaking Discoveries of Patricia Goldman-Rakic 1937 - 2003Dokument13 SeitenFeature Article The Neurobiology of Thought: The Groundbreaking Discoveries of Patricia Goldman-Rakic 1937 - 2003Adelina MarițiuNoch keine Bewertungen
- Nihms 588714Dokument31 SeitenNihms 588714Diana BastoNoch keine Bewertungen
- Affective Neuroscience and PsychopathlogyDokument78 SeitenAffective Neuroscience and PsychopathlogyLucíaNoch keine Bewertungen
- McDonald-Messinger Empathy DevelopmentDokument36 SeitenMcDonald-Messinger Empathy DevelopmentMihaela CiubotaruNoch keine Bewertungen
- MOANS - Trail-Making Test, Stroop, and COWATDokument50 SeitenMOANS - Trail-Making Test, Stroop, and COWATGeorge BialkowskiNoch keine Bewertungen
- CVS TreatmentDokument6 SeitenCVS TreatmentNaguibYunosNoch keine Bewertungen
- Phgy 556 Course Outline 2018 PreliminaryDokument7 SeitenPhgy 556 Course Outline 2018 PreliminaryAnonymous t6S5qw2glPNoch keine Bewertungen
- The Gender You Are and The Gender You Like - Sexual Preference and Empathic Neural ResponsesDokument10 SeitenThe Gender You Are and The Gender You Like - Sexual Preference and Empathic Neural Responsesgabriela marmolejoNoch keine Bewertungen
- PTH Migraine LoopDokument15 SeitenPTH Migraine LoopRenju KuriakoseNoch keine Bewertungen
- Brodman MapDokument18 SeitenBrodman MapBecky Edwards0% (1)
- Brain Teen GuideDokument45 SeitenBrain Teen GuideTacha Von LammNoch keine Bewertungen
- DecenteringDokument14 SeitenDecenteringLv MarkNoch keine Bewertungen
- Schizophr Bull 1994 Gordon 697 712Dokument16 SeitenSchizophr Bull 1994 Gordon 697 712EdwardVargasNoch keine Bewertungen
- Cerebral Hemispheres 2018 PDFDokument69 SeitenCerebral Hemispheres 2018 PDFlailatul husnaNoch keine Bewertungen
- 02 Neurobiologia de La AdiccionDokument25 Seiten02 Neurobiologia de La AdiccionZaira Camila CriolloNoch keine Bewertungen
- Biological Aspects of OCD SeminarDokument60 SeitenBiological Aspects of OCD Seminarneha mattikoppaNoch keine Bewertungen
- Social Neuroscience Key ReadingsDokument335 SeitenSocial Neuroscience Key ReadingsMartin Herrera100% (2)
- The Age of Magical Overthinking: Notes on Modern IrrationalityVon EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityBewertung: 4 von 5 Sternen4/5 (24)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisVon EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisBewertung: 4.5 von 5 Sternen4.5/5 (42)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsVon EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNoch keine Bewertungen
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedVon EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedBewertung: 5 von 5 Sternen5/5 (80)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsVon EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsBewertung: 5 von 5 Sternen5/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaVon EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaBewertung: 4.5 von 5 Sternen4.5/5 (266)