Das könnte Ihnen auch gefallen
- Reporte Ing GenéticaDokument8 SeitenReporte Ing GenéticaMike BurtonNoch keine Bewertungen
- Transcripcion de Eucariotas PDFDokument31 SeitenTranscripcion de Eucariotas PDFEduardo Mata GonzálezNoch keine Bewertungen
- PRACTICA NO1 Descripcion de Material de LaboratorioDokument12 SeitenPRACTICA NO1 Descripcion de Material de LaboratorioYolanda Jael Rizo FonsecaNoch keine Bewertungen
- Ejercicio Señales 2 FisiologiaDokument3 SeitenEjercicio Señales 2 FisiologiaYoali Asiain LeconaNoch keine Bewertungen
- 1 Operon de LactosaDokument4 Seiten1 Operon de Lactosafrancisco lozano gomezNoch keine Bewertungen
- Preparación MTTDokument20 SeitenPreparación MTTJuan Camilo JuradoNoch keine Bewertungen
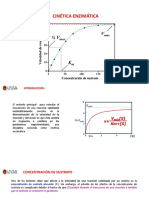
- Tema 8 - Cinetica Enzimatica Farmacia PDFDokument12 SeitenTema 8 - Cinetica Enzimatica Farmacia PDFMariciell Larissa Gonzales TiconaNoch keine Bewertungen
- Experimento 8 Lisozima 06-04Dokument20 SeitenExperimento 8 Lisozima 06-04rominagriseldaNoch keine Bewertungen
- Estabilidad Del RNAmDokument31 SeitenEstabilidad Del RNAmEduardo Mata GonzálezNoch keine Bewertungen
- DNA PlasmidicoDokument3 SeitenDNA PlasmidicoAbraham GuzmanNoch keine Bewertungen
- Manual de Analitica III 2 de MarzoDokument56 SeitenManual de Analitica III 2 de MarzoCarlos AyonNoch keine Bewertungen
- 1 Valorar La Solución de NaOH Con Ftalato Ácido de PotasioDokument3 Seiten1 Valorar La Solución de NaOH Con Ftalato Ácido de PotasioYamilka Yanneth Solís MojicaNoch keine Bewertungen
- Reporte 1. Equipo 6Dokument9 SeitenReporte 1. Equipo 6LiLiana NiñoNoch keine Bewertungen
- Ejercicions No Mendel Agosto 2020Dokument2 SeitenEjercicions No Mendel Agosto 2020ellectrocompulsivoNoch keine Bewertungen
- Clase Practica 26Dokument5 SeitenClase Practica 26Kenya SantacruzNoch keine Bewertungen
- Ing. EnzimáticaDokument46 SeitenIng. EnzimáticaArmando Montoya García100% (1)
- Analisis e Interpretacion de InsertosDokument10 SeitenAnalisis e Interpretacion de InsertosRichard Dennïs NinaNoch keine Bewertungen
- Citoesqueleto ForoDokument39 SeitenCitoesqueleto ForoClaudia Montes GuerraNoch keine Bewertungen
- Ejercicios de SecuencuacionDokument2 SeitenEjercicios de Secuencuacionhernando guerrero0% (1)
- Cuadro Medios de Cultivo-AmbientalDokument3 SeitenCuadro Medios de Cultivo-AmbientalYordy Rodriguez100% (1)
- Enzimas y Coenzimas, PDFDokument7 SeitenEnzimas y Coenzimas, PDFdharmaNoch keine Bewertungen
- Regulación Genética en El Operón Lac Corregido 1Dokument4 SeitenRegulación Genética en El Operón Lac Corregido 1Miguel RivasNoch keine Bewertungen
- Autoevaluacion TisularDokument6 SeitenAutoevaluacion TisularCabito CatalanNoch keine Bewertungen
- Receptores CelularesDokument36 SeitenReceptores CelularesyovanaNoch keine Bewertungen
- CofactoresDokument3 SeitenCofactoresmaryprafloNoch keine Bewertungen
- Cuadro Comparativo Replicación 2Dokument2 SeitenCuadro Comparativo Replicación 2Eswin Octavio Minera LemusNoch keine Bewertungen
- EpibatidinaDokument8 SeitenEpibatidinaMonchis Lizett HernandezNoch keine Bewertungen
- Chaperonas y ChaperoninasDokument26 SeitenChaperonas y ChaperoninasaarongondraNoch keine Bewertungen
- Reporte Practica 1Dokument9 SeitenReporte Practica 1Carlos Cerón LinaresNoch keine Bewertungen
- Práctica 5 Extraccion de ADN PlasmidicoDokument6 SeitenPráctica 5 Extraccion de ADN PlasmidicoFranklin Joel Eche PoloNoch keine Bewertungen
- Apuntes y Tablas Técnicas EspectroscópicasDokument16 SeitenApuntes y Tablas Técnicas EspectroscópicasjulioNoch keine Bewertungen
- Presentacion 6 Cinetica EnzimaticaDokument46 SeitenPresentacion 6 Cinetica EnzimaticaE Clinton LMNoch keine Bewertungen
- Cromatografia para Purificación de ProteínasDokument55 SeitenCromatografia para Purificación de ProteínasAlejandro VieyraNoch keine Bewertungen
- Informe 11 - PCR Reacción en Cadena de La PolimerasaDokument5 SeitenInforme 11 - PCR Reacción en Cadena de La PolimerasaJuan Jose Rendon OchoaNoch keine Bewertungen
- ELECTROFORESISDokument14 SeitenELECTROFORESISCristian RecaldeNoch keine Bewertungen
- Generalidades FEUMDokument1 SeiteGeneralidades FEUMcarlosNoch keine Bewertungen
- Extraccion Adn VegetalDokument4 SeitenExtraccion Adn Vegetaljose davidNoch keine Bewertungen
- Clase 1 2019IIDokument59 SeitenClase 1 2019IIKatty Julieth Rangel GutierrezNoch keine Bewertungen
- DiastereómerosDokument7 SeitenDiastereómerosAndrea ReynosoNoch keine Bewertungen
- Cuestionario Práctica 6 - Técnicas de Tinción DiferencialesDokument4 SeitenCuestionario Práctica 6 - Técnicas de Tinción DiferencialesLizzet SalazarNoch keine Bewertungen
- Problemas Finales de CCDokument10 SeitenProblemas Finales de CCJaza VlogsNoch keine Bewertungen
- Cromatografia y ColesterolDokument3 SeitenCromatografia y ColesterolGonzález Chávez MarianaNoch keine Bewertungen
- Practica FinalDokument3 SeitenPractica FinalMILENKA YOMAR MONTEALEGRE SARZURINoch keine Bewertungen
- Taller Transcripción DNA Octubre - 2020Dokument4 SeitenTaller Transcripción DNA Octubre - 2020licy villanuevaNoch keine Bewertungen
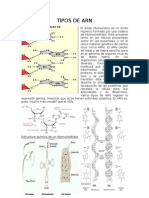
- Tipos de ArnDokument5 SeitenTipos de ArnMiguel Angel Rodas Herrera100% (2)
- Diseño Del Sistema de Seguridad y Salud en El Trabajo en El Área Del Almacén en El Laboratorio de BioprocesosDokument49 SeitenDiseño Del Sistema de Seguridad y Salud en El Trabajo en El Área Del Almacén en El Laboratorio de BioprocesosElianis Fonte GonzalezNoch keine Bewertungen
- Aislamiento de La CaseinaDokument9 SeitenAislamiento de La Caseinajerson chafloque ruizNoch keine Bewertungen
- 5 ElectroforesisDokument84 Seiten5 ElectroforesisDiana RiosNoch keine Bewertungen
- Práctica 3 - Aislamiento de LectinasDokument2 SeitenPráctica 3 - Aislamiento de LectinasDaniela IbarraNoch keine Bewertungen
- Aislamiento de Adn PlasmídicoDokument6 SeitenAislamiento de Adn PlasmídicoGus VedderNoch keine Bewertungen
- Diseño de PrimersDokument6 SeitenDiseño de PrimersAdriana Herrera VivasNoch keine Bewertungen
- Uniones 1Dokument1 SeiteUniones 1Sofía Torroledo RamírezNoch keine Bewertungen
- 06 Laboratorio N°6 Extracción y Determinación de ColesterolDokument3 Seiten06 Laboratorio N°6 Extracción y Determinación de ColesterolMacarena Fernanda LizamaNoch keine Bewertungen
- Tarea CitoesqueletoDokument6 SeitenTarea CitoesqueletoSebastián Rodríguez VelásquezNoch keine Bewertungen
- Practica 1 Fisicoquimica III TerminadaDokument11 SeitenPractica 1 Fisicoquimica III TerminadaRoss HernandezNoch keine Bewertungen
- Reporte 6Dokument12 SeitenReporte 6JUAN DIEGO VALDEZ QUISPENoch keine Bewertungen
- Tema 2 ReplicacionDokument25 SeitenTema 2 ReplicacionArsenio Flores VasquezNoch keine Bewertungen
- ReplicacionDokument28 SeitenReplicacionAurelia Gonzales CalleNoch keine Bewertungen
- Semana13 Teoria ReplicacionADNDokument21 SeitenSemana13 Teoria ReplicacionADNCARMEN LINDA SUAREZNoch keine Bewertungen
- Replicación, Transcripción y Traducción.Dokument4 SeitenReplicación, Transcripción y Traducción.Valentina VergaraNoch keine Bewertungen
- Información General AmtDokument4 SeitenInformación General AmtPABLO GRANJANoch keine Bewertungen
- Plan de Saguridad Defensa CivilDokument24 SeitenPlan de Saguridad Defensa CivilAlvaro JosuéNoch keine Bewertungen
- AUDITORIA O EVALUACION DE LA GESTION DEL TALENTO HUMANO Tarea 8Dokument7 SeitenAUDITORIA O EVALUACION DE LA GESTION DEL TALENTO HUMANO Tarea 8geri100% (1)
- Exposición de Alfombras Antiguas Españolas (1933) PDFDokument53 SeitenExposición de Alfombras Antiguas Españolas (1933) PDFBeatriz ÁlvarezNoch keine Bewertungen
- Guía 8° y Ticket Comprensión Lectora 13Dokument2 SeitenGuía 8° y Ticket Comprensión Lectora 13CristalNoch keine Bewertungen
- Propuesta Proyecto de PalmaDokument7 SeitenPropuesta Proyecto de PalmaJonathan LaguadoNoch keine Bewertungen
- Marco General y Proceso Constructivo Del Puente Allccomachay-1-25Dokument25 SeitenMarco General y Proceso Constructivo Del Puente Allccomachay-1-25Angelo Renato Lazo ParedesNoch keine Bewertungen
- 19 20CinematicaIIDokument2 Seiten19 20CinematicaIIAdriana BioscaNoch keine Bewertungen
- Canalón Autoportante ABM 680 PDFDokument1 SeiteCanalón Autoportante ABM 680 PDFJavier HerreraNoch keine Bewertungen
- 6 Ast Tecnica-Indice Jose Antonio AguileraDokument4 Seiten6 Ast Tecnica-Indice Jose Antonio AguileraGirey pahuanaNoch keine Bewertungen
- 4 Operaciones Con Monomios IIDokument23 Seiten4 Operaciones Con Monomios IIAna CuentalNoch keine Bewertungen
- Mol530 s1 Estrategia Competitiva PorterDokument30 SeitenMol530 s1 Estrategia Competitiva PorterJosselin Aguirre GarnicaNoch keine Bewertungen
- Estados Financieros ProyectadosDokument23 SeitenEstados Financieros ProyectadosRaul Arenas AsteteNoch keine Bewertungen
- Problemario Resumen Estadístico - 2im49Dokument14 SeitenProblemario Resumen Estadístico - 2im49Sara LaurenesNoch keine Bewertungen
- Evaluaciones 4to Periodo 3ero y 4toDokument29 SeitenEvaluaciones 4to Periodo 3ero y 4tosirleyNoch keine Bewertungen
- Formato No 2 - Plan de TrabajoDokument3 SeitenFormato No 2 - Plan de TrabajosdsdsNoch keine Bewertungen
- Teoria Del Final en La Partida de Ajedrez - Ganzo (1957) PDFDokument94 SeitenTeoria Del Final en La Partida de Ajedrez - Ganzo (1957) PDFanfichoelosNoch keine Bewertungen
- Equipo6 D5E4 ForEmDokument2 SeitenEquipo6 D5E4 ForEmÁngel DavidNoch keine Bewertungen
- Cuanto Falta para El 5 de Enero - Buscar Con GoogleDokument1 SeiteCuanto Falta para El 5 de Enero - Buscar Con Googleqpnpdz8kpxNoch keine Bewertungen
- Caso BancafeDokument11 SeitenCaso BancafeRogelio RodriguezNoch keine Bewertungen
- Lab09 - Variador de Velocidad ACDokument34 SeitenLab09 - Variador de Velocidad ACCesar Herrera100% (1)
- Psicotécnico CemDokument27 SeitenPsicotécnico Cemcesar davidNoch keine Bewertungen
- MARCO TEORICO Terminal-1Dokument22 SeitenMARCO TEORICO Terminal-1Marylena GutierrezNoch keine Bewertungen
- Alma BotanicaDokument17 SeitenAlma BotanicaBetianaNoch keine Bewertungen
- Proyecto PatagoniaDokument11 SeitenProyecto PatagoniaClaudio CascoNoch keine Bewertungen
- Prostatitis Aguda y CronicaDokument17 SeitenProstatitis Aguda y CronicaTati M SuxeNoch keine Bewertungen
- Ups CT005203Dokument128 SeitenUps CT005203Juan Carlos FajardoNoch keine Bewertungen
- Aase LRPD FinalDokument3 SeitenAase LRPD FinalMaría Aburto VásquezNoch keine Bewertungen
- Diferencias Entre Liderazgo y GestionDokument2 SeitenDiferencias Entre Liderazgo y GestionezarauzNoch keine Bewertungen
- Trabajo Final para ImprimirDokument50 SeitenTrabajo Final para ImprimirXIMENA GALARZANoch keine Bewertungen