Das könnte Ihnen auch gefallen
- Stiff-Person Syndrome and Related Disorders: A Comprehensive, Practical GuideVon EverandStiff-Person Syndrome and Related Disorders: A Comprehensive, Practical GuideNoch keine Bewertungen
- Gen SchizopDokument11 SeitenGen SchizopDewiakyuNoch keine Bewertungen
- MR Findings in Leigh Syndrome With COX Deficiency and SURF-1 MutationsDokument6 SeitenMR Findings in Leigh Syndrome With COX Deficiency and SURF-1 MutationsDrsandy SandyNoch keine Bewertungen
- tmp1329 TMPDokument11 Seitentmp1329 TMPFrontiersNoch keine Bewertungen
- Synaptic Dysbindin-1 Reductions in Schizophrenia Occur in An Isoform-Specific Manner Indicating Their Subsynaptic LocationDokument14 SeitenSynaptic Dysbindin-1 Reductions in Schizophrenia Occur in An Isoform-Specific Manner Indicating Their Subsynaptic LocationCinthyaMárquezRojasNoch keine Bewertungen
- Cei 13095Dokument6 SeitenCei 13095karen rinconNoch keine Bewertungen
- 1 s2.0 S016517811530559X MainDokument6 Seiten1 s2.0 S016517811530559X MainDiana DionisioNoch keine Bewertungen
- SchizophreniaDokument6 SeitenSchizophreniaМ.К. МариNoch keine Bewertungen
- BIO 199 HandoutDokument5 SeitenBIO 199 HandoutChristian CustodioNoch keine Bewertungen
- Martinez2016 PDFDokument12 SeitenMartinez2016 PDFPedroFigueroaNoch keine Bewertungen
- Braak Hipotesis For ParkinsonDokument22 SeitenBraak Hipotesis For Parkinsonfirza yoga baskoroNoch keine Bewertungen
- Srinivasan 2016Dokument16 SeitenSrinivasan 2016Zeljko TomljanovicNoch keine Bewertungen
- Acetylsistein Journal 1Dokument13 SeitenAcetylsistein Journal 1Arifa LainaNoch keine Bewertungen
- Microduplication of xp22.31Dokument4 SeitenMicroduplication of xp22.31Roberth SanchezNoch keine Bewertungen
- Gim 201618Dokument8 SeitenGim 201618Ioana PoterasNoch keine Bewertungen
- Pereira MSDokument10 SeitenPereira MSAntónio AlvimNoch keine Bewertungen
- Human-Leukocyte Antigen Class II Genes in Earlyonset Obsessive-Compulsive DisorderDokument8 SeitenHuman-Leukocyte Antigen Class II Genes in Earlyonset Obsessive-Compulsive DisorderBerenice RomeroNoch keine Bewertungen
- Post Mortem Brain Study AutismDokument8 SeitenPost Mortem Brain Study Autism5bs48gjk94Noch keine Bewertungen
- Genotype-Phenotype Studies in Three Families PQBP1Dokument9 SeitenGenotype-Phenotype Studies in Three Families PQBP1Mosabbira RahmanNoch keine Bewertungen
- Expression of Zonula Occludens-1 (ZO-1) and The Transcription Factor ZO-1-associated Nucleic Acid-Binding Protei..Dokument16 SeitenExpression of Zonula Occludens-1 (ZO-1) and The Transcription Factor ZO-1-associated Nucleic Acid-Binding Protei..juanNoch keine Bewertungen
- Paper 1Dokument11 SeitenPaper 1Anonymous K8fT6HNoch keine Bewertungen
- J Jpsychires 2015 12 013Dokument7 SeitenJ Jpsychires 2015 12 013Fitri anaNoch keine Bewertungen
- Pagano 2016Dokument29 SeitenPagano 2016Naruto UzumakiNoch keine Bewertungen
- MiceDokument10 SeitenMicedavid.brieberNoch keine Bewertungen
- Poster Abstracts: Biological PsychiatryDokument1 SeitePoster Abstracts: Biological PsychiatryafsdsaadsNoch keine Bewertungen
- Meta-Analysis Shows Strong Positive AssociationDokument8 SeitenMeta-Analysis Shows Strong Positive AssociationFahrunnisa NurdinNoch keine Bewertungen
- Jurnal HirschsprungDokument6 SeitenJurnal HirschsprungVicky RizalNoch keine Bewertungen
- Is Brain-Derived Neurotrophic Factor (BDNF) Val66Met Polymorphism Associated With Obsessive-Compulsive Disorder? A Meta-AnalysisDokument7 SeitenIs Brain-Derived Neurotrophic Factor (BDNF) Val66Met Polymorphism Associated With Obsessive-Compulsive Disorder? A Meta-AnalysisSylvie jNoch keine Bewertungen
- LINE-1 Hypomethylation Is Associated With Poor Risperidone Response in A First Episode of Psychosis CohortDokument12 SeitenLINE-1 Hypomethylation Is Associated With Poor Risperidone Response in A First Episode of Psychosis CohortDiogo MarquesNoch keine Bewertungen
- Deriziotis-2014-De Novo TBR1 MutatioDokument12 SeitenDeriziotis-2014-De Novo TBR1 Mutatiopriyaseshadri5Noch keine Bewertungen
- FTPDokument325 SeitenFTPKim Chwin KhyeNoch keine Bewertungen
- Hayashi Takagi 2014Dokument9 SeitenHayashi Takagi 2014Petros SkapinakisNoch keine Bewertungen
- SIMI - Volume Comunicazioni Orali e PosterDokument212 SeitenSIMI - Volume Comunicazioni Orali e Postermaria wahyuniNoch keine Bewertungen
- LupusDokument7 SeitenLupusZyanCeronNoch keine Bewertungen
- The Molecular Pathology of Schizophrenia: An Overview of Existing Knowledge and New Directions For Future ResearchDokument22 SeitenThe Molecular Pathology of Schizophrenia: An Overview of Existing Knowledge and New Directions For Future ResearchJorge SalazarNoch keine Bewertungen
- Frontotemporal Dementia NeuropathologyDokument14 SeitenFrontotemporal Dementia Neuropathologyppico1963Noch keine Bewertungen
- The Pathophysiology and Genetics of OCDDokument12 SeitenThe Pathophysiology and Genetics of OCDCrescent FangNoch keine Bewertungen
- Journal Pre-Proof: Biological PsychiatryDokument36 SeitenJournal Pre-Proof: Biological PsychiatryJon hahaNoch keine Bewertungen
- 2018 Oh LabMedOnlineDokument4 Seiten2018 Oh LabMedOnlineEduardo PasteneNoch keine Bewertungen
- гени-превод СараDokument5 Seitenгени-превод СараNikola VeleskiNoch keine Bewertungen
- PlosOne 2011Dokument8 SeitenPlosOne 2011Annetrude de MooijNoch keine Bewertungen
- Jakobsson 2015Dokument9 SeitenJakobsson 2015Priscila Navarro MedinaNoch keine Bewertungen
- Dysbindin-1 Involvement in The Etiology of Schizophrenia: ReviewDokument14 SeitenDysbindin-1 Involvement in The Etiology of Schizophrenia: ReviewAlicia Carrera TorresNoch keine Bewertungen
- Genetics and AlzheimersDokument15 SeitenGenetics and AlzheimersSex & Gender Women's Health CollaborativeNoch keine Bewertungen
- The Disruption of Profiling of Serotonergic Neurons, Results in Autism-Related BehaviorsDokument22 SeitenThe Disruption of Profiling of Serotonergic Neurons, Results in Autism-Related BehaviorsAngelinni Taglioni StangeNoch keine Bewertungen
- Solapamiento de Genes Entre Toc y EsqDokument7 SeitenSolapamiento de Genes Entre Toc y EsqLucas MartínNoch keine Bewertungen
- Genetic Overlap Between Autism, Schizophrenia and Bipolar DisorderDokument7 SeitenGenetic Overlap Between Autism, Schizophrenia and Bipolar DisorderamarillonoexpectaNoch keine Bewertungen
- 2016.gouel Et AlDokument5 Seiten2016.gouel Et AlBruce DO VANNoch keine Bewertungen
- Fpsyt-12-655178 37Dokument10 SeitenFpsyt-12-655178 37Fitri anaNoch keine Bewertungen
- Liver X receptor β regulates the development of the dentate gyrus and autistic-like behavior in the mouseDokument9 SeitenLiver X receptor β regulates the development of the dentate gyrus and autistic-like behavior in the mouseDárcio CasteloNoch keine Bewertungen
- 62 - Searching For The Immune Basis of Obsessive-Compulsive DisorderDokument7 Seiten62 - Searching For The Immune Basis of Obsessive-Compulsive DisorderPaula CantalapiedraNoch keine Bewertungen
- 10 1002/ajmg B 30713Dokument3 Seiten10 1002/ajmg B 30713Sri HariNoch keine Bewertungen
- Gene and SchizophreniaDokument22 SeitenGene and SchizophreniaaldensmagdalenaNoch keine Bewertungen
- 1 s2.0 S266645932300046X MainDokument10 Seiten1 s2.0 S266645932300046X MainEmmanuel OwonaNoch keine Bewertungen
- HHS Public Access: Studying Polyglutamine Diseases in DrosophilaDokument39 SeitenHHS Public Access: Studying Polyglutamine Diseases in DrosophilaJoeNoch keine Bewertungen
- Nature 2014-11 Wen DISC1Dokument26 SeitenNature 2014-11 Wen DISC1mark175511Noch keine Bewertungen
- Expressional Control of The Constitutive' Isoforms of Nitric Oxide Synthase (NOS I and NOS III)Dokument18 SeitenExpressional Control of The Constitutive' Isoforms of Nitric Oxide Synthase (NOS I and NOS III)Ravi TejaNoch keine Bewertungen
- 2Dokument9 Seiten2Mostafa FakhriNoch keine Bewertungen
- Gut Microbiota Is Altered in Patients With Alzheimer's DiseaseDokument10 SeitenGut Microbiota Is Altered in Patients With Alzheimer's DiseaseaedrNoch keine Bewertungen
- TMP A6 B9Dokument12 SeitenTMP A6 B9FrontiersNoch keine Bewertungen
- MIlosz, Czeslaw - To Begin Where I Am (2001, FSG) PDFDokument481 SeitenMIlosz, Czeslaw - To Begin Where I Am (2001, FSG) PDFRavenna100% (6)
- Cassidy2013 Attachment Theory and Research PDFDokument39 SeitenCassidy2013 Attachment Theory and Research PDFRavenna0% (1)
- HAZMAT Happens: The Premiere Hazmat Training For The Healthcare ProfessionalDokument2 SeitenHAZMAT Happens: The Premiere Hazmat Training For The Healthcare ProfessionalRavennaNoch keine Bewertungen
- HHS Public Access: Hoarding Disorder: More Than Just A Problem of Too Much StuffDokument5 SeitenHHS Public Access: Hoarding Disorder: More Than Just A Problem of Too Much StuffRavennaNoch keine Bewertungen
- Stenglerwenzke2007 PDFDokument8 SeitenStenglerwenzke2007 PDFRavennaNoch keine Bewertungen
- HHS Public Access: Antidepressant Use and Cognitive Decline: The Health and Retirement StudyDokument16 SeitenHHS Public Access: Antidepressant Use and Cognitive Decline: The Health and Retirement StudyRavennaNoch keine Bewertungen
- 10 1016@j Jocn 2018 10 042Dokument11 Seiten10 1016@j Jocn 2018 10 042RavennaNoch keine Bewertungen
- Is A Psychodynamic Perspective Relevant To The Clinical Management of Obsessive-Compulsive Disorder?Dokument12 SeitenIs A Psychodynamic Perspective Relevant To The Clinical Management of Obsessive-Compulsive Disorder?RavennaNoch keine Bewertungen
- Role of Psilocybin in The Treatment of Depression: Ananya Mahapatra and Rishi GuptaDokument3 SeitenRole of Psilocybin in The Treatment of Depression: Ananya Mahapatra and Rishi GuptaRavennaNoch keine Bewertungen
- Immune Aberrations in Obsessive-Compulsive Disorder: A Systematic Review and Meta-AnalysisDokument9 SeitenImmune Aberrations in Obsessive-Compulsive Disorder: A Systematic Review and Meta-AnalysisRavennaNoch keine Bewertungen
- Bokor 2014Dokument15 SeitenBokor 2014RavennaNoch keine Bewertungen
- Neurotransmitters, Temperament and Social Functioning: ReviewDokument14 SeitenNeurotransmitters, Temperament and Social Functioning: ReviewRavennaNoch keine Bewertungen
- A Case Series of 223 Patients With Depersonalization-Derealization SyndromeDokument11 SeitenA Case Series of 223 Patients With Depersonalization-Derealization SyndromeRavennaNoch keine Bewertungen
- Ostracism and Indirect Reciprocity: The Reproductive Significance of HumorDokument18 SeitenOstracism and Indirect Reciprocity: The Reproductive Significance of HumorRavennaNoch keine Bewertungen
- N-Acetylcysteine in The Treatment of Psychiatric Disorders: Current Status and Future ProspectsDokument51 SeitenN-Acetylcysteine in The Treatment of Psychiatric Disorders: Current Status and Future ProspectsRavennaNoch keine Bewertungen
- Nihms836647 PDFDokument4 SeitenNihms836647 PDFRavennaNoch keine Bewertungen
- Personality Disorders and Culture: Contemporary Clinical Views (Part B)Dokument13 SeitenPersonality Disorders and Culture: Contemporary Clinical Views (Part B)RavennaNoch keine Bewertungen
- Furey2006 Antidepressant ScopolamineDokument9 SeitenFurey2006 Antidepressant ScopolamineRavennaNoch keine Bewertungen
- Umr Cnrs 5249, Grenoble, France Cea, DSV, Irtsv, Grenoble, France Àuniversite Joseph Fourier, Grenoble, FranceDokument10 SeitenUmr Cnrs 5249, Grenoble, France Cea, DSV, Irtsv, Grenoble, France Àuniversite Joseph Fourier, Grenoble, FranceRavennaNoch keine Bewertungen
- Radiologi Intervensi Di IndiaDokument3 SeitenRadiologi Intervensi Di IndiaPutu Adi SusantaNoch keine Bewertungen
- 24.18.00 - Chikungunya2 DR RAGADokument48 Seiten24.18.00 - Chikungunya2 DR RAGAjcvh24Noch keine Bewertungen
- Neonatal SepsisDokument6 SeitenNeonatal SepsisSunaina AdhikariNoch keine Bewertungen
- SaveBooklet BookletDokument120 SeitenSaveBooklet BookletRoscelie KhoNoch keine Bewertungen
- Wound HealingDokument70 SeitenWound HealingSachin KpNoch keine Bewertungen
- Emergency Care (Hemophilia)Dokument30 SeitenEmergency Care (Hemophilia)Nica Georgelle Maniego SamonteNoch keine Bewertungen
- CellCommunication Assessment3Dokument6 SeitenCellCommunication Assessment3Michaela Maria GarciaNoch keine Bewertungen
- TaxonomyDokument71 SeitenTaxonomyNabeel Uddin100% (1)
- Pat An AnewDokument126 SeitenPat An AnewgshchurovskiyNoch keine Bewertungen
- Notice: Grants and Cooperative Agreements Availability, Etc.: Women Living in Puerto Rico and U.S. Virgin Islands HIV Prevention ProgramDokument8 SeitenNotice: Grants and Cooperative Agreements Availability, Etc.: Women Living in Puerto Rico and U.S. Virgin Islands HIV Prevention ProgramJustia.comNoch keine Bewertungen
- Post Graduate ClinicsDokument113 SeitenPost Graduate ClinicsSnehal PatilNoch keine Bewertungen
- MVG09Dokument4 SeitenMVG09Sondos HarbiehNoch keine Bewertungen
- AUB - Physical Examination of UrineDokument3 SeitenAUB - Physical Examination of UrineJeanne RodiñoNoch keine Bewertungen
- One Health ConceptDokument2 SeitenOne Health ConceptYume AkamaiNoch keine Bewertungen
- Personal StatementDokument1 SeitePersonal StatementTaiwo Oyeniyi100% (2)
- 2022 Field Research Book DAD - 1190 - 28250Dokument262 Seiten2022 Field Research Book DAD - 1190 - 28250Diego PalhariniNoch keine Bewertungen
- Dyslipidemia: Darmono SSDokument35 SeitenDyslipidemia: Darmono SSLa Ode Rinaldi0% (1)
- 2020 11 07 XII Economics 1Dokument17 Seiten2020 11 07 XII Economics 1Sjft6hd FfNoch keine Bewertungen
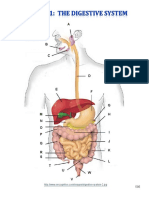
- Digestive System Charts 1Dokument14 SeitenDigestive System Charts 1api-306762527Noch keine Bewertungen
- Phlegmon: by - Dharmendra Singh Group No. - 317Dokument15 SeitenPhlegmon: by - Dharmendra Singh Group No. - 317Mouna Jomaa DhifaouiNoch keine Bewertungen
- Anestesi EpiduralDokument2 SeitenAnestesi EpiduralShazean ShahNoch keine Bewertungen
- MIDTERMSDokument24 SeitenMIDTERMSCherish Marie HurbodaNoch keine Bewertungen
- AFA ReviwerDokument21 SeitenAFA ReviwerLovely day ybanezNoch keine Bewertungen
- The Importance of SleepDokument4 SeitenThe Importance of Sleep7A323Trọng NhânNoch keine Bewertungen
- CHAPTER 4: Modifiable Risk Factors of Lifestyle Diseases: SmokingDokument4 SeitenCHAPTER 4: Modifiable Risk Factors of Lifestyle Diseases: SmokingShimah100% (2)
- 91931-Article Text-260086-1-10-20131218Dokument8 Seiten91931-Article Text-260086-1-10-20131218Liana OpriţăNoch keine Bewertungen
- Glossary of Commonly Used Dental Terms: 14212 Ambaum Boulevard Southwest Suite #101, Burien, WA 98166Dokument10 SeitenGlossary of Commonly Used Dental Terms: 14212 Ambaum Boulevard Southwest Suite #101, Burien, WA 98166Mayra RojasNoch keine Bewertungen
- Stroke:: What Should Nurses Need To Know About Stroke?Dokument68 SeitenStroke:: What Should Nurses Need To Know About Stroke?Suci Aning TNoch keine Bewertungen
- SyncytiumDokument87 SeitenSyncytiumjustin sNoch keine Bewertungen
- Self-Love Affirmations For Deep Sleep: Raise self-worth Build confidence, Heal your wounded heart, Reprogram your subconscious mind, 8-hour sleep cycle, know your value, effortless healingsVon EverandSelf-Love Affirmations For Deep Sleep: Raise self-worth Build confidence, Heal your wounded heart, Reprogram your subconscious mind, 8-hour sleep cycle, know your value, effortless healingsBewertung: 4.5 von 5 Sternen4.5/5 (6)
- Allen Carr's Easy Way to Quit Vaping: Get Free from JUUL, IQOS, Disposables, Tanks or any other Nicotine ProductVon EverandAllen Carr's Easy Way to Quit Vaping: Get Free from JUUL, IQOS, Disposables, Tanks or any other Nicotine ProductBewertung: 5 von 5 Sternen5/5 (31)
- Healing Your Aloneness: Finding Love and Wholeness Through Your Inner ChildVon EverandHealing Your Aloneness: Finding Love and Wholeness Through Your Inner ChildBewertung: 3.5 von 5 Sternen3.5/5 (9)
- Breaking Addiction: A 7-Step Handbook for Ending Any AddictionVon EverandBreaking Addiction: A 7-Step Handbook for Ending Any AddictionBewertung: 4.5 von 5 Sternen4.5/5 (2)
- Allen Carr's Easy Way to Quit Smoking Without Willpower: The best-selling quit smoking method updated for the 21st centuryVon EverandAllen Carr's Easy Way to Quit Smoking Without Willpower: The best-selling quit smoking method updated for the 21st centuryBewertung: 5 von 5 Sternen5/5 (47)
- Alcoholics Anonymous, Fourth Edition: The official "Big Book" from Alcoholic AnonymousVon EverandAlcoholics Anonymous, Fourth Edition: The official "Big Book" from Alcoholic AnonymousBewertung: 5 von 5 Sternen5/5 (22)
- Save Me from Myself: How I Found God, Quit Korn, Kicked Drugs, and Lived to Tell My StoryVon EverandSave Me from Myself: How I Found God, Quit Korn, Kicked Drugs, and Lived to Tell My StoryNoch keine Bewertungen
- Guts: The Endless Follies and Tiny Triumphs of a Giant DisasterVon EverandGuts: The Endless Follies and Tiny Triumphs of a Giant DisasterBewertung: 4 von 5 Sternen4/5 (99)
- Quitting Smoking & Vaping For Dummies: 2nd EditionVon EverandQuitting Smoking & Vaping For Dummies: 2nd EditionNoch keine Bewertungen
- Little Red Book: Alcoholics AnonymousVon EverandLittle Red Book: Alcoholics AnonymousBewertung: 5 von 5 Sternen5/5 (20)
- Easyway Express: Stop Smoking and Quit E-CigarettesVon EverandEasyway Express: Stop Smoking and Quit E-CigarettesBewertung: 5 von 5 Sternen5/5 (15)
- Allen Carr's Quit Drinking Without Willpower: Be a happy nondrinkerVon EverandAllen Carr's Quit Drinking Without Willpower: Be a happy nondrinkerBewertung: 5 von 5 Sternen5/5 (8)
- The Heart of Addiction: A New Approach to Understanding and Managing Alcoholism and Other Addictive BehaviorsVon EverandThe Heart of Addiction: A New Approach to Understanding and Managing Alcoholism and Other Addictive BehaviorsNoch keine Bewertungen
- Twelve Steps and Twelve Traditions: The “Twelve and Twelve” — Essential Alcoholics Anonymous readingVon EverandTwelve Steps and Twelve Traditions: The “Twelve and Twelve” — Essential Alcoholics Anonymous readingBewertung: 5 von 5 Sternen5/5 (11)
- Stop Drinking Now: The original Easyway methodVon EverandStop Drinking Now: The original Easyway methodBewertung: 5 von 5 Sternen5/5 (29)
- Living Sober: Practical methods alcoholics have used for living without drinkingVon EverandLiving Sober: Practical methods alcoholics have used for living without drinkingBewertung: 4 von 5 Sternen4/5 (50)
- The Easy Way to Stop Gambling: Take Control of Your LifeVon EverandThe Easy Way to Stop Gambling: Take Control of Your LifeBewertung: 4 von 5 Sternen4/5 (197)
- You Are More Than This Will Ever Be: Methamphetamine: The Dirty DrugVon EverandYou Are More Than This Will Ever Be: Methamphetamine: The Dirty DrugNoch keine Bewertungen
- THE FRUIT YOU’LL NEVER SEE: A memoir about overcoming shame.Von EverandTHE FRUIT YOU’LL NEVER SEE: A memoir about overcoming shame.Bewertung: 4 von 5 Sternen4/5 (7)
- Healing the Scars of Addiction: Reclaiming Your Life and Moving into a Healthy FutureVon EverandHealing the Scars of Addiction: Reclaiming Your Life and Moving into a Healthy FutureBewertung: 4.5 von 5 Sternen4.5/5 (7)
- Blood Orange Night: My Journey to the Edge of MadnessVon EverandBlood Orange Night: My Journey to the Edge of MadnessBewertung: 4 von 5 Sternen4/5 (42)
- The Stop Drinking Expert: Alcohol Lied to Me Updated And Extended EditionVon EverandThe Stop Drinking Expert: Alcohol Lied to Me Updated And Extended EditionBewertung: 5 von 5 Sternen5/5 (63)
- 12 Step Spirituality: Every Person’s Guide to Taking the Twelve StepsVon Everand12 Step Spirituality: Every Person’s Guide to Taking the Twelve StepsBewertung: 5 von 5 Sternen5/5 (17)
- The Kindness Method: Change Your Habits for Good Using Self-Compassion and UnderstandingVon EverandThe Kindness Method: Change Your Habits for Good Using Self-Compassion and UnderstandingBewertung: 5 von 5 Sternen5/5 (10)
- Trailer Park Parable: A Memoir of How Three Brothers Strove to Rise Above Their Broken Past, Find Forgiveness, and Forge a Hopeful FutureVon EverandTrailer Park Parable: A Memoir of How Three Brothers Strove to Rise Above Their Broken Past, Find Forgiveness, and Forge a Hopeful FutureBewertung: 4.5 von 5 Sternen4.5/5 (6)
- Alcoholics Anonymous: The Landmark of Recovery and Vital LivingVon EverandAlcoholics Anonymous: The Landmark of Recovery and Vital LivingBewertung: 4.5 von 5 Sternen4.5/5 (291)