Das könnte Ihnen auch gefallen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Dysrhythmias: Normal Electrical ActivityDokument32 SeitenDysrhythmias: Normal Electrical ActivityLarisse de LeonNoch keine Bewertungen
- Pex 09 06Dokument4 SeitenPex 09 06Illich Ramirez Tanta100% (2)
- Meat Milk and Fish HygieneDokument74 SeitenMeat Milk and Fish Hygieneransingh100% (1)
- Sensory & Motor Tracts of Spinal ChordDokument38 SeitenSensory & Motor Tracts of Spinal ChordtiiandiNoch keine Bewertungen
- Court of The Minor ClansDokument59 SeitenCourt of The Minor Clansdipstick789Noch keine Bewertungen
- Lecture 2-Aviation Human Factor (Flight Physiology)Dokument47 SeitenLecture 2-Aviation Human Factor (Flight Physiology)Richard Thodé Jr100% (1)
- Common Medical AbbriviationDokument13 SeitenCommon Medical AbbriviationADAMU LAWALNoch keine Bewertungen
- Acute Phase ProteinsDokument3 SeitenAcute Phase ProteinswaihenyaNoch keine Bewertungen
- Occlusal AnalysisDokument14 SeitenOcclusal AnalysisDavid DongNoch keine Bewertungen
- Animal NutritionDokument6 SeitenAnimal NutritionMeily ZoelNoch keine Bewertungen
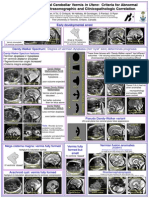
- Abnormal VermisDokument1 SeiteAbnormal VermiscalmlandNoch keine Bewertungen
- LeprosyDokument38 SeitenLeprosym6000000Noch keine Bewertungen
- Work Sheet 5 The Skin A28 Group1Dokument2 SeitenWork Sheet 5 The Skin A28 Group1Japet Floyd AlipioNoch keine Bewertungen
- For The Strength of The Pack Is The Wolf, and The Strength of The Wolf Is The Pack.Dokument10 SeitenFor The Strength of The Pack Is The Wolf, and The Strength of The Wolf Is The Pack.Phone Myat Pyaye SoneNoch keine Bewertungen
- MaternityDokument91 SeitenMaternityAnonymous D8KswoNoch keine Bewertungen
- Ace Reasoning New AddaDokument359 SeitenAce Reasoning New Addajuinarka24112002Noch keine Bewertungen
- Acute AbdomenDokument4 SeitenAcute Abdomensarguss14Noch keine Bewertungen
- Advanced Guide To Chakra and Charts V17Dokument34 SeitenAdvanced Guide To Chakra and Charts V17Magia Del Mar100% (2)
- Biography of Anton Van LeeuwenhoekDokument7 SeitenBiography of Anton Van LeeuwenhoekDevul KhandelwalNoch keine Bewertungen
- Digestive System AnatomyDokument8 SeitenDigestive System AnatomyBrittany Lei MaquirayaNoch keine Bewertungen
- Data Review Meeting Template 2018 Final 1Dokument19 SeitenData Review Meeting Template 2018 Final 1AdraNoch keine Bewertungen
- Pollen Allergy - Prevention and Natural Remedies - Greater KashmirDokument3 SeitenPollen Allergy - Prevention and Natural Remedies - Greater Kashmirrizan loanNoch keine Bewertungen
- Quiz ReproductiveDokument78 SeitenQuiz ReproductiveMedShare86% (7)
- GuppyDokument2 SeitenGuppymartynunez12345Noch keine Bewertungen
- Ielts 5.0 l1 R DoneDokument14 SeitenIelts 5.0 l1 R DoneMai TrangNoch keine Bewertungen
- Surgery CssDokument13 SeitenSurgery CssNaren ShanNoch keine Bewertungen
- A Clinical ExemplarDokument5 SeitenA Clinical Exemplarapi-302128044100% (1)
- Pyeronie Disease and HomoeopathyDokument16 SeitenPyeronie Disease and HomoeopathyDr. Rajneesh Kumar Sharma MD HomNoch keine Bewertungen
- Risk Management Prelim ReviewerDokument3 SeitenRisk Management Prelim ReviewerCharmaigne BañaresNoch keine Bewertungen
- CABERGOLINE Safety2Dokument6 SeitenCABERGOLINE Safety2Kane SmithNoch keine Bewertungen