Das könnte Ihnen auch gefallen
- UF1811 - Manejo de equipos de medida de contaminantes atmosféricosVon EverandUF1811 - Manejo de equipos de medida de contaminantes atmosféricosNoch keine Bewertungen
- Celulas Eucariotas, Procariotas y Archaeas ComparaciónDokument4 SeitenCelulas Eucariotas, Procariotas y Archaeas ComparaciónCatherine RangelNoch keine Bewertungen
- BiomoléculasArias PDFDokument584 SeitenBiomoléculasArias PDFAndrea Morales100% (1)
- Protones en LevaduraDokument7 SeitenProtones en Levaduralauraj5Noch keine Bewertungen
- Estequiometria MicrobianaDokument3 SeitenEstequiometria MicrobianaArianaNoch keine Bewertungen
- Expo Organica ResonanciaDokument14 SeitenExpo Organica ResonanciaAlex Douglas Suarez RuizNoch keine Bewertungen
- La Importancia de La Química en La MicrobiologíaDokument5 SeitenLa Importancia de La Química en La MicrobiologíaBrian Ogawa100% (1)
- Química Orgánica - McMurry - Cengage - 9° EdiciónDokument2 SeitenQuímica Orgánica - McMurry - Cengage - 9° EdiciónLuz Tapia Villarroel0% (2)
- PIRROLDokument7 SeitenPIRROLArnold Alfredo TumiNoch keine Bewertungen
- Revista de Divulgación - Mutagenesis Dirigida AplicacionesDokument6 SeitenRevista de Divulgación - Mutagenesis Dirigida AplicacionesKeren Espinoza100% (1)
- Taller Eteres, Tioles, EpoxidosDokument2 SeitenTaller Eteres, Tioles, EpoxidosAnonymous GdWMlV46bUNoch keine Bewertungen
- Aplicación Industrial Del AnisolDokument1 SeiteAplicación Industrial Del AnisolNigu100% (1)
- Bioquímica Lehninger CAPITULO 01Dokument21 SeitenBioquímica Lehninger CAPITULO 01Lyd Granados75% (8)
- Práctica Nº5 Propiedades Molares ParcialesDokument7 SeitenPráctica Nº5 Propiedades Molares ParcialesRomel RauraNoch keine Bewertungen
- Ejercicio 1Dokument3 SeitenEjercicio 1Nayelli Cervantes100% (2)
- BLASTDokument34 SeitenBLASTJesús SaucedoNoch keine Bewertungen
- Examen Cinetica QuimicaDokument9 SeitenExamen Cinetica QuimicaPedro Reynaldo Marin Dominguez100% (1)
- Programa de Biociencias Médicas IDokument4 SeitenPrograma de Biociencias Médicas IEdwin TapiasNoch keine Bewertungen
- Acetales y HemiacetalesDokument13 SeitenAcetales y HemiacetalesKazukiRonnyLiverpool100% (14)
- Practica 7Dokument9 SeitenPractica 7Gabo LiceaNoch keine Bewertungen
- Activación de Los Ácidos GrasosDokument2 SeitenActivación de Los Ácidos GrasosJaviera Gotuzzo CarrascoNoch keine Bewertungen
- Guía para El Examen de Conocimientos. Instituto de Biotecnologia UNAMDokument13 SeitenGuía para El Examen de Conocimientos. Instituto de Biotecnologia UNAMCarlos Alberto Ramirez Garcia50% (2)
- Preguntas Molecular Segundo CorteDokument6 SeitenPreguntas Molecular Segundo CorteValentina Arango100% (1)
- Bio Energetic A Torre de ElectronesDokument18 SeitenBio Energetic A Torre de ElectronesMadeleyne Rodriguez50% (2)
- Dominios de La VidaDokument4 SeitenDominios de La VidaGissela MéndezNoch keine Bewertungen
- Hidrolisis Acida y Enzimatica Del AlmidonDokument6 SeitenHidrolisis Acida y Enzimatica Del AlmidonGilberto Diaz Medina100% (1)
- Infografía Tipos de Medios de CultivoDokument1 SeiteInfografía Tipos de Medios de CultivoFlores Huescas Adriana Abril100% (1)
- Fracción Másica y Fracción Molar-Fórmulas y EjemplosDokument2 SeitenFracción Másica y Fracción Molar-Fórmulas y EjemplosEstefany CabreraNoch keine Bewertungen
- Mapa MetabólicoDokument3 SeitenMapa MetabólicoBarbs Vn0% (1)
- 5.2 Transcripcion de La Informacion Del ADNDokument4 Seiten5.2 Transcripcion de La Informacion Del ADNDaniela AlvaradoNoch keine Bewertungen
- Clase 4 - Estequiometría - 2020Dokument37 SeitenClase 4 - Estequiometría - 2020Sol Moniec100% (1)
- Qué Es Química Analítica y Su ClasificaciónDokument3 SeitenQué Es Química Analítica y Su ClasificaciónAndreita Sanchez100% (2)
- Derivados de Acidos CarboxilicosDokument6 SeitenDerivados de Acidos CarboxilicossiriusdarkNoch keine Bewertungen
- Biosíntesis-2Dokument59 SeitenBiosíntesis-2Maleja Matoma100% (1)
- Precipitación SalinaDokument5 SeitenPrecipitación SalinaOrlando Baca0% (1)
- Práctica de Laboratorio # 2 - Preparación de Un Hidrogel de Magnetita-Alginato para La Remoción de Un ColoranteDokument3 SeitenPráctica de Laboratorio # 2 - Preparación de Un Hidrogel de Magnetita-Alginato para La Remoción de Un ColoranteBautista PV100% (1)
- ProblemarioDokument10 SeitenProblemariomagNoch keine Bewertungen
- Metabolismo MicrobianoDokument5 SeitenMetabolismo MicrobianoPatricia Sanabria0% (1)
- Cuestionario de Morfologia BacterianaDokument6 SeitenCuestionario de Morfologia BacterianaSaulvarNoch keine Bewertungen
- Primer Parcial de Botánica OpcionalDokument14 SeitenPrimer Parcial de Botánica OpcionalJenny Amparo Gutierrez MartinezNoch keine Bewertungen
- Hongos OrnamentalesDokument4 SeitenHongos OrnamentalesmauricioNoch keine Bewertungen
- Calibracion Multi Variable Modelo Directo Imprtante-1Dokument8 SeitenCalibracion Multi Variable Modelo Directo Imprtante-1Emily Coromoto Gonzalez MarcanoNoch keine Bewertungen
- Asignación 13. Tipos y Medidas de Cajas para La Cosecha de Frutas y Hortalizas PDFDokument5 SeitenAsignación 13. Tipos y Medidas de Cajas para La Cosecha de Frutas y Hortalizas PDFMónica INoch keine Bewertungen
- Taller Parejas 1 SN EDokument2 SeitenTaller Parejas 1 SN EOmar Jose Morales CameloNoch keine Bewertungen
- Clase 2 Estructura AtomicaDokument128 SeitenClase 2 Estructura AtomicaDaniela Sierra Quiroz50% (2)
- Deterioro de LipidosDokument3 SeitenDeterioro de LipidosEmely Bustamante RojasNoch keine Bewertungen
- Tinciones DiferencialesDokument6 SeitenTinciones Diferencialesalvaro acNoch keine Bewertungen
- Determinación de Carbonatos y BicarbonatosDokument4 SeitenDeterminación de Carbonatos y BicarbonatosRomario GutierrezNoch keine Bewertungen
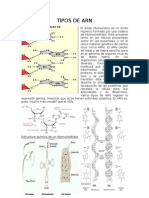
- Tipos de ArnDokument5 SeitenTipos de ArnMiguel Angel Rodas Herrera100% (2)
- CARACTERIZACIÓN DEL BIORREACTOR Bioflo 200Dokument8 SeitenCARACTERIZACIÓN DEL BIORREACTOR Bioflo 20013marielNoch keine Bewertungen
- Preinforme Aldehidos y CetonasDokument4 SeitenPreinforme Aldehidos y CetonasSantiago Cifuentes Almanza100% (1)
- Doc1 2Dokument33 SeitenDoc1 2Betzabet Montalvo VillegasNoch keine Bewertungen
- Estructura Molecular de Los Ácidos NucleicosDokument3 SeitenEstructura Molecular de Los Ácidos NucleicosleschaltelNoch keine Bewertungen
- Estructura Molecular de Los Ácidos NucleicosDokument2 SeitenEstructura Molecular de Los Ácidos NucleicosDaniela Franco HNoch keine Bewertungen
- Traducción Del Trabajo Original de Watson y CrickDokument3 SeitenTraducción Del Trabajo Original de Watson y CrickSergio SánchezNoch keine Bewertungen
- Estructura Molecular de Los Ácidos Nucleicos PDFDokument3 SeitenEstructura Molecular de Los Ácidos Nucleicos PDFRichieQCNoch keine Bewertungen
- Traduccion Articulo ADNDokument3 SeitenTraduccion Articulo ADNEsmeralda Angeles GarciaNoch keine Bewertungen
- Estructura Molecular de Los Ácidos NucleicosDokument3 SeitenEstructura Molecular de Los Ácidos NucleicosJuan Esteban Sánchez bernalNoch keine Bewertungen
- Deseamos Proponer Una Estructura para La Sal Del Ácido Nucleico de DesoxirribosaDokument4 SeitenDeseamos Proponer Una Estructura para La Sal Del Ácido Nucleico de Desoxirribosaidali velazquezNoch keine Bewertungen
- 4 Taller Nucleotidos y Acidos NucleicosDokument19 Seiten4 Taller Nucleotidos y Acidos NucleicosSSergio Andres Santamaria TorresNoch keine Bewertungen
- INTA - CRCordoba - EEAManfredi - Grandon N. - Puesta - A - Punto - Del - Metodo - de - Extraccion - de - Adn - Con - PDFDokument1 SeiteINTA - CRCordoba - EEAManfredi - Grandon N. - Puesta - A - Punto - Del - Metodo - de - Extraccion - de - Adn - Con - PDFManu RodriguezNoch keine Bewertungen
- Minipreps, Tipos de Biotecnologia y Tipos de PromotoresDokument12 SeitenMinipreps, Tipos de Biotecnologia y Tipos de PromotoresManu RodriguezNoch keine Bewertungen
- Minipreps, Tipos de Biotecnologia y Tipos de PromotoresDokument12 SeitenMinipreps, Tipos de Biotecnologia y Tipos de PromotoresManu RodriguezNoch keine Bewertungen
- Manual de Prácticas de Bioinformática - (PG 1 - 45)Dokument45 SeitenManual de Prácticas de Bioinformática - (PG 1 - 45)Manu Rodriguez75% (8)
- Seminario 10B - Terapia Génica PDFDokument14 SeitenSeminario 10B - Terapia Génica PDFBrisa NarvaezNoch keine Bewertungen
- Memorias 2012Dokument455 SeitenMemorias 2012Manu RodriguezNoch keine Bewertungen
- Generación Del Repertorio de AnticuerposDokument2 SeitenGeneración Del Repertorio de AnticuerposManu RodriguezNoch keine Bewertungen
- Guia de Bk2017Dokument45 SeitenGuia de Bk2017Manu RodriguezNoch keine Bewertungen
- Maduración de Linfocitos B y Reordenamiento de IgsDokument1 SeiteMaduración de Linfocitos B y Reordenamiento de IgsManu RodriguezNoch keine Bewertungen
- Mecánica de Rocas Pucp Parte 1Dokument73 SeitenMecánica de Rocas Pucp Parte 1Piero H VizcarraNoch keine Bewertungen
- Evolución en La Legislación Ambiental en MéxicoDokument6 SeitenEvolución en La Legislación Ambiental en MéxicoElsy RodherzNoch keine Bewertungen
- Alicia CamaraDokument13 SeitenAlicia CamaraMalena MastricchioNoch keine Bewertungen
- Tabla Del Tiempo GeológicoDokument3 SeitenTabla Del Tiempo GeológicoIrvin Villanueva Nieto100% (1)
- Informe 3 - Elaboracion de Tablas de VidaDokument4 SeitenInforme 3 - Elaboracion de Tablas de Vidafede100% (1)
- 100 Preguntas de Geologia y MineralogiaDokument2 Seiten100 Preguntas de Geologia y MineralogiaSantiago Parra JimenezNoch keine Bewertungen
- Turner KayapoDokument24 SeitenTurner KayapoIsrael OrtizNoch keine Bewertungen
- Ficha Didáctica Nº2 Reino Hongo Protoctista 2º Inaes 2019Dokument7 SeitenFicha Didáctica Nº2 Reino Hongo Protoctista 2º Inaes 2019Isabel BarriosNoch keine Bewertungen
- Frustraciones InfantilesDokument4 SeitenFrustraciones InfantilesGeorgina Aquino GarciaNoch keine Bewertungen
- Plancton, Bentos y NectonDokument3 SeitenPlancton, Bentos y NectonRube Kisai Sanchez CuatecontziNoch keine Bewertungen
- Capitulo 1. - Comodidad Térmica. Curso de Confort Térmico Andino-PunoDokument23 SeitenCapitulo 1. - Comodidad Térmica. Curso de Confort Térmico Andino-PunofredyNoch keine Bewertungen
- 2.guia Seleccion Multiple HdeCDokument1 Seite2.guia Seleccion Multiple HdeCMJ_CJ_NINoch keine Bewertungen
- Eddea Arquitectura y UrbanismoDokument4 SeitenEddea Arquitectura y UrbanismoMartin FeNoch keine Bewertungen
- Proyección de La VozDokument16 SeitenProyección de La Vozlucia simosa50% (2)
- Tirak L 500 PDokument172 SeitenTirak L 500 PDanielNoch keine Bewertungen
- INGENIERIA DE DETALLE Parte 4Dokument20 SeitenINGENIERIA DE DETALLE Parte 4Marysabel SanchezNoch keine Bewertungen
- 9 Analisis de Alimentos ClaseDokument44 Seiten9 Analisis de Alimentos Clasescribdmortis0% (2)
- Triptico TsunamiDokument4 SeitenTriptico TsunamiMia67% (3)
- Libro I y II Del Régimen Del Suelo - Pugs FinalDokument160 SeitenLibro I y II Del Régimen Del Suelo - Pugs FinalMiguel Rosero Haro100% (2)
- Manifiesto 3 - Nikola Tesla, Calentamiento Global y VacunasDokument10 SeitenManifiesto 3 - Nikola Tesla, Calentamiento Global y VacunasJose Antonio Ortiz RodriguezNoch keine Bewertungen
- Firmas Expectrales Aplicado A GeologiaDokument3 SeitenFirmas Expectrales Aplicado A GeologiaHr McNoch keine Bewertungen
- PAPER (Guayas y Arenamiento)Dokument12 SeitenPAPER (Guayas y Arenamiento)mariannysNoch keine Bewertungen
- Estudio Hidrologico GoyllarisquizgaDokument77 SeitenEstudio Hidrologico GoyllarisquizgaJUkarevi VicenteNoch keine Bewertungen
- QUIMICADokument20 SeitenQUIMICAmaria lucia moreno100% (1)
- Teorias 1Dokument5 SeitenTeorias 1Blanca PerezNoch keine Bewertungen
- Enfermedad Profesional y Su Aseguramiento NicaraguaDokument14 SeitenEnfermedad Profesional y Su Aseguramiento NicaraguaAngela Sanchez100% (1)
- Analisis Sismico en El Departamento Del CesarDokument47 SeitenAnalisis Sismico en El Departamento Del CesarElias Ernesto RojasNoch keine Bewertungen
- Como Enseñar La QuimicaDokument29 SeitenComo Enseñar La QuimicaJhonny Moreno100% (1)
- Capitulo 16 17 18Dokument3 SeitenCapitulo 16 17 18MERCEDESNoch keine Bewertungen
- Las MezclasDokument3 SeitenLas MezclasMaria Jose Ocaranza IriarteNoch keine Bewertungen