Das könnte Ihnen auch gefallen
- Oogenesis and ovarian histology of the South American lizard Salvator merianaeDokument14 SeitenOogenesis and ovarian histology of the South American lizard Salvator merianaeValeria Garcia ValdezNoch keine Bewertungen
- Nakanishi 2018Dokument63 SeitenNakanishi 2018sorbakti sinagaNoch keine Bewertungen
- Zelfstudie Van Blastula Naar 3 KiemlagenDokument18 SeitenZelfstudie Van Blastula Naar 3 KiemlagenCheyenne ProvoostNoch keine Bewertungen
- Crozatier Et Al-2007-Cellular MicrobiologyDokument10 SeitenCrozatier Et Al-2007-Cellular MicrobiologyVINEET KUREELNoch keine Bewertungen
- Canine Fetus Immune System at Late DevelopmentDokument4 SeitenCanine Fetus Immune System at Late DevelopmentWillNoch keine Bewertungen
- Biol 233 Uofc Review PackageDokument15 SeitenBiol 233 Uofc Review PackagePeter KimNoch keine Bewertungen
- ANIMAL SYMMETRY AND TISSUE GRADEDokument6 SeitenANIMAL SYMMETRY AND TISSUE GRADERajdeep GhoshNoch keine Bewertungen
- Post-Natal Development of Prescapular Lymph Node in One-Humped Camel (Camelus Dromedarius) - A Histological StudyDokument12 SeitenPost-Natal Development of Prescapular Lymph Node in One-Humped Camel (Camelus Dromedarius) - A Histological StudyFrancisco SantosNoch keine Bewertungen
- TonsilasDokument15 SeitenTonsilasLuz InfanteNoch keine Bewertungen
- Auxinas - 2a ClaseDokument7 SeitenAuxinas - 2a ClaseBrayan JiménezNoch keine Bewertungen
- WJG 6 169 PDFDokument8 SeitenWJG 6 169 PDFJaldo FreireNoch keine Bewertungen
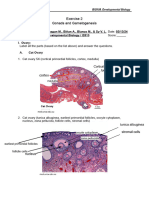
- Final Exercise 2Dokument20 SeitenFinal Exercise 2myca.blancoNoch keine Bewertungen
- The Fundamental Unit of The Life - CELLDokument13 SeitenThe Fundamental Unit of The Life - CELLPralex PrajapatiNoch keine Bewertungen
- Histology of Umbilical Cord in MammalsDokument16 SeitenHistology of Umbilical Cord in MammalsFia NisaNoch keine Bewertungen
- Morphofunction Development of The White Rat Thymus in Postnatal OntogenesisDokument5 SeitenMorphofunction Development of The White Rat Thymus in Postnatal OntogenesisCentral Asian StudiesNoch keine Bewertungen
- The peripheral vesicles gather multivesicular bodies with different behaviorDokument11 SeitenThe peripheral vesicles gather multivesicular bodies with different behaviorUNIG Aluana SantanaNoch keine Bewertungen
- Embryology GastrulationDokument9 SeitenEmbryology GastrulationRivera Gómez América LucíaNoch keine Bewertungen
- Alveolar RegenerationDokument12 SeitenAlveolar RegenerationWidia MeliawatiNoch keine Bewertungen
- The Immune System of Amphibians: An OverviewDokument7 SeitenThe Immune System of Amphibians: An OverviewandreNoch keine Bewertungen
- Laporan Lisa Selsai Edit LampranDokument22 SeitenLaporan Lisa Selsai Edit LampranAnalianahsNoch keine Bewertungen
- Ls1 Final Study GuideDokument7 SeitenLs1 Final Study GuideMinh NguyenNoch keine Bewertungen
- Biology S.S.S. 2 FIRST TERM SCHEME of WorkDokument16 SeitenBiology S.S.S. 2 FIRST TERM SCHEME of WorkLeoverseNoch keine Bewertungen
- Human Body OrganizationDokument6 SeitenHuman Body OrganizationWolverineInZenNoch keine Bewertungen
- Buchtova 2007 Zoology II PDFDokument21 SeitenBuchtova 2007 Zoology II PDFEduardo Panadero CuarteroNoch keine Bewertungen
- Developmental BiologyDokument6 SeitenDevelopmental BiologyBismarck John SalazarNoch keine Bewertungen
- Medical Parasitology Lecture L400Dokument68 SeitenMedical Parasitology Lecture L400emmanuel imahNoch keine Bewertungen
- Elife 43828Dokument19 SeitenElife 43828jaypasco79Noch keine Bewertungen
- Bio 101 General BiologyDokument7 SeitenBio 101 General BiologyMohammed kashimNoch keine Bewertungen
- Camp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.Von EverandCamp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.Noch keine Bewertungen
- Human EmbriologyDokument67 SeitenHuman EmbriologyGalang Perdamaian0% (1)
- Cells and Development: MorphogenesisDokument13 SeitenCells and Development: Morphogenesisosandi wijayasooriyaNoch keine Bewertungen
- Phylogeny and ontogeny of the immune systemDokument5 SeitenPhylogeny and ontogeny of the immune systemGaurav KumarNoch keine Bewertungen
- Ellismar - 1802101010003 - Kelas 01Dokument17 SeitenEllismar - 1802101010003 - Kelas 01EllismarNoch keine Bewertungen
- The Phylum CtenophoraDokument3 SeitenThe Phylum CtenophoraTI Journals PublishingNoch keine Bewertungen
- Phylum CtenophoraDokument23 SeitenPhylum CtenophoramsnidanusratNoch keine Bewertungen
- Platyhelminthes and MolluscaDokument7 SeitenPlatyhelminthes and MolluscaIsnaeni RachmawatiNoch keine Bewertungen
- ArthropodsDokument6 SeitenArthropodsGintang PrayogiNoch keine Bewertungen
- Pudney 1995 PDFDokument39 SeitenPudney 1995 PDFKei MlNoch keine Bewertungen
- Nilsson 2017Dokument18 SeitenNilsson 2017نذير الشرعبيNoch keine Bewertungen
- Baroli Leite Et Al (2015) Comparative Testis Morphology of Neotropical AnuransDokument10 SeitenBaroli Leite Et Al (2015) Comparative Testis Morphology of Neotropical AnuransJuan Carlos SánchezNoch keine Bewertungen
- Paternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inDokument27 SeitenPaternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inじょしら フィアンナNoch keine Bewertungen
- Six Major Steps in Animal Evolution: Are We Derived Sponge Larvae?Dokument17 SeitenSix Major Steps in Animal Evolution: Are We Derived Sponge Larvae?cypriano68100% (2)
- Sed 202 Biological Science Research Tasks BDokument8 SeitenSed 202 Biological Science Research Tasks BBryllie BautistaNoch keine Bewertungen
- Gao Zhang 2018 Chapter Amphi OxusDokument44 SeitenGao Zhang 2018 Chapter Amphi OxusAnil KumarNoch keine Bewertungen
- Spermatogonial Stem Cells: Questions, Models and PerspectivesDokument8 SeitenSpermatogonial Stem Cells: Questions, Models and PerspectivesKsenia KamenskaiaNoch keine Bewertungen
- AnatomyDokument16 SeitenAnatomywakhalewakhaleNoch keine Bewertungen
- Biogenesis and Cell TheoryDokument6 SeitenBiogenesis and Cell TheoryScarlet OrigenesNoch keine Bewertungen
- HO1 - 8 - Biology April 23-24 PDFDokument10 SeitenHO1 - 8 - Biology April 23-24 PDFSaghana SaravanakarthikNoch keine Bewertungen
- Fish reproduction organs and developmentDokument11 SeitenFish reproduction organs and developmentsksingl350Noch keine Bewertungen
- Alten Develop 2012Dokument10 SeitenAlten Develop 2012zbc7z9x0tNoch keine Bewertungen
- FungiDokument31 SeitenFungiTADIWANASHE TINONETSANANoch keine Bewertungen
- PART B GRADE 12 PRACTICAL SPOTTING (1)Dokument5 SeitenPART B GRADE 12 PRACTICAL SPOTTING (1)cdsingh74Noch keine Bewertungen
- NHLBI Workshop Summary: The Mysterious Pulmonary Brush CellDokument4 SeitenNHLBI Workshop Summary: The Mysterious Pulmonary Brush CellcholoconvertiNoch keine Bewertungen
- CESTODE - PPT MajorDokument92 SeitenCESTODE - PPT MajorJOSEPH NDERITU100% (2)
- ProtozoaDokument25 SeitenProtozoanupur.kmcNoch keine Bewertungen
- AxolotlsDokument15 SeitenAxolotlsagathapavlovaNoch keine Bewertungen
- 2723NEETZoologyGrowth RepairandAgeingTheory - UnlockedDokument9 Seiten2723NEETZoologyGrowth RepairandAgeingTheory - UnlockedJayaraju BattulaNoch keine Bewertungen
- Human Embryonic DevelopmentDokument8 SeitenHuman Embryonic DevelopmentGargi BhattacharyaNoch keine Bewertungen
- REPRO - HISTOWORldDokument7 SeitenREPRO - HISTOWORldKarl Torres Uganiza RmtNoch keine Bewertungen
- There Is No Doubt ThatDokument7 SeitenThere Is No Doubt ThatAhmad Beni RoufNoch keine Bewertungen
- Zhou2014 PDFDokument7 SeitenZhou2014 PDFAhmad Beni RoufNoch keine Bewertungen
- Reed2014 PDFDokument13 SeitenReed2014 PDFAhmad Beni RoufNoch keine Bewertungen
- I09214649 PDFDokument4 SeitenI09214649 PDFAhmad Beni RoufNoch keine Bewertungen
- Solomon and Udoji PDFDokument14 SeitenSolomon and Udoji PDFAhmad Beni RoufNoch keine Bewertungen
- Rayto RT 2100 C Microplate Reader Technical SpecsDokument1 SeiteRayto RT 2100 C Microplate Reader Technical SpecsAhmad Beni RoufNoch keine Bewertungen
- Mitigating Intracohort Cannibalism in Cultured Freshwater FishDokument16 SeitenMitigating Intracohort Cannibalism in Cultured Freshwater FishAhmad Beni RoufNoch keine Bewertungen
- Jurnal Ilmiah Mahasiswa Pendidikan Biologi, Volume 1, Issue 1, Agustus 2016, Hal 95-106Dokument12 SeitenJurnal Ilmiah Mahasiswa Pendidikan Biologi, Volume 1, Issue 1, Agustus 2016, Hal 95-106Ahmad Beni RoufNoch keine Bewertungen
- Viral Vaccine For Farmed FishDokument17 SeitenViral Vaccine For Farmed FishAhmad Beni RoufNoch keine Bewertungen
- Brosur ToyoboDokument8 SeitenBrosur ToyobodanilriosNoch keine Bewertungen
- Political Science Assignment Sem v-1Dokument4 SeitenPolitical Science Assignment Sem v-1Aayush SinhaNoch keine Bewertungen
- Magsaysay v. NLRCDokument17 SeitenMagsaysay v. NLRCJohn BernalNoch keine Bewertungen
- UntitledDokument1.422 SeitenUntitledKarinNoch keine Bewertungen
- Literary Alexandria PDFDokument50 SeitenLiterary Alexandria PDFMariaFrankNoch keine Bewertungen
- Benito Mussolini - English (Auto-Generated)Dokument35 SeitenBenito Mussolini - English (Auto-Generated)FJ MacaleNoch keine Bewertungen
- 2023 Whole Bible Reading PlanDokument2 Seiten2023 Whole Bible Reading PlanDenmark BulanNoch keine Bewertungen
- Immigration and Exploration of The USADokument6 SeitenImmigration and Exploration of The USAТетяна МешкоNoch keine Bewertungen
- The Problem and It'S BackgroundDokument9 SeitenThe Problem and It'S BackgroundGisbert Martin LayaNoch keine Bewertungen
- Bacaan Bilal Shalat Tarawih 11 Rakaat Bahasa IndonesiaDokument4 SeitenBacaan Bilal Shalat Tarawih 11 Rakaat Bahasa IndonesiaMTSs Tahfizh Ulumul Quran MedanNoch keine Bewertungen
- List of applicants for admission to Non-UT Pool categoryDokument102 SeitenList of applicants for admission to Non-UT Pool categoryvishalNoch keine Bewertungen
- Self Concept's Role in Buying BehaviorDokument6 SeitenSelf Concept's Role in Buying BehaviorMadhavi GundabattulaNoch keine Bewertungen
- Procter &gamble: Our PurposeDokument39 SeitenProcter &gamble: Our Purposemubasharabdali5373Noch keine Bewertungen
- Gramatyka Test 14Dokument1 SeiteGramatyka Test 14lolmem100% (1)
- Delhi Public School Mid-Term Exam for Class 5 Students Covering English, Math, ScienceDokument20 SeitenDelhi Public School Mid-Term Exam for Class 5 Students Covering English, Math, ScienceFaaz MohammadNoch keine Bewertungen
- CH8 OligopolyModelsDokument37 SeitenCH8 OligopolyModelsJeffrey LaoNoch keine Bewertungen
- Universiti Kuala Lumpur Intra Management System: Unikl-ImsDokument16 SeitenUniversiti Kuala Lumpur Intra Management System: Unikl-ImsAfnanFadlanBinAzmiNoch keine Bewertungen
- Annex D Initial Evaluation Results IER 2Dokument6 SeitenAnnex D Initial Evaluation Results IER 2ruffaNoch keine Bewertungen
- A Gateway For SIP Event Interworking: - Sasu Tarkoma & Thalainayar Balasubramanian RamyaDokument20 SeitenA Gateway For SIP Event Interworking: - Sasu Tarkoma & Thalainayar Balasubramanian RamyaMadhunath YadavNoch keine Bewertungen
- CalendarDokument1 SeiteCalendarapi-277854872Noch keine Bewertungen
- Mutual Fund: Advnatages and Disadvantages To InvestorsDokument11 SeitenMutual Fund: Advnatages and Disadvantages To InvestorsSnehi GuptaNoch keine Bewertungen
- P&WC S.B. No. A1427R3Dokument16 SeitenP&WC S.B. No. A1427R3Elmer Villegas100% (1)
- Organized Crime in Central America The Northern Triangle, Report On The Americas #29 DRAFTDokument125 SeitenOrganized Crime in Central America The Northern Triangle, Report On The Americas #29 DRAFTLa GringaNoch keine Bewertungen
- Reflection Paper AristotleDokument3 SeitenReflection Paper AristotleMelissa Sullivan67% (9)
- EH2741 HT16 Lecture 5 PDFDokument25 SeitenEH2741 HT16 Lecture 5 PDFNiels Romanovich RomanovNoch keine Bewertungen
- Resume With Leadership New 2Dokument2 SeitenResume With Leadership New 2api-356808363Noch keine Bewertungen
- 2021 Pslce Selection - ShedDokument254 Seiten2021 Pslce Selection - ShedMolley KachingweNoch keine Bewertungen
- Understanding Plane and Solid FiguresDokument9 SeitenUnderstanding Plane and Solid FiguresCoronia Mermaly Lamsen100% (2)
- The Canterbury Tales Is A Collection of Stories Written in Middle English byDokument25 SeitenThe Canterbury Tales Is A Collection of Stories Written in Middle English byyoonglespianoNoch keine Bewertungen
- IVAN's TEST UAS 1Dokument9 SeitenIVAN's TEST UAS 1gilangNoch keine Bewertungen
- Problem 7 - 16Dokument2 SeitenProblem 7 - 16Jao FloresNoch keine Bewertungen