Das könnte Ihnen auch gefallen
- Expo de FisiologiaDokument65 SeitenExpo de Fisiologiaabi100% (1)
- Resumen SINAPSISDokument10 SeitenResumen SINAPSISEsme Monroy0% (1)
- Clase 3Dokument11 SeitenClase 3danielNoch keine Bewertungen
- Unidad 1 - Clase 3 - Potencial de Reposo - Potencial de Acción - Transmisión SinápticaDokument36 SeitenUnidad 1 - Clase 3 - Potencial de Reposo - Potencial de Acción - Transmisión SinápticaAntonia Samur ContrerasNoch keine Bewertungen
- SinapsisDokument5 SeitenSinapsisYamila RamirezNoch keine Bewertungen
- Apuntes Fisio 2-2Dokument6 SeitenApuntes Fisio 2-2Arnau Miquel CosNoch keine Bewertungen
- Neuro SinapsisDokument5 SeitenNeuro SinapsisTatiana EspinoNoch keine Bewertungen
- NeurotransmisoresDokument18 SeitenNeurotransmisoresAgustina GuzmánNoch keine Bewertungen
- Tema 8-Bloque II-Comunicacion Neuronal SinapsisDokument11 SeitenTema 8-Bloque II-Comunicacion Neuronal SinapsisEdwin Macas MorenoNoch keine Bewertungen
- Tema 3 - 2 SinapsisDokument65 SeitenTema 3 - 2 SinapsisGreysmar Vanessa Chirinos GuapeNoch keine Bewertungen
- Potenciales de Receptor o Sensorial 1rtryDokument81 SeitenPotenciales de Receptor o Sensorial 1rtryMichael100% (2)
- NeurofisiollgíaDokument3 SeitenNeurofisiollgíaAgostina EsmorisNoch keine Bewertungen
- Clase Comunicacion Celular 2019-1Dokument36 SeitenClase Comunicacion Celular 2019-1RebecaNoch keine Bewertungen
- Tema 4. Transmisión SinápticaDokument9 SeitenTema 4. Transmisión SinápticaJose García SegoviaNoch keine Bewertungen
- Tema 13 NeurotrasmisoresDokument7 SeitenTema 13 NeurotrasmisoresFrancisco PabonNoch keine Bewertungen
- Sinapsis: La unidad funcional del sistema nerviosoDokument5 SeitenSinapsis: La unidad funcional del sistema nerviosoBrandon GonzalezNoch keine Bewertungen
- NeurotransmisoresDokument16 SeitenNeurotransmisoresEstefany MorenoNoch keine Bewertungen
- TEMA 4 Completo + LibroDokument10 SeitenTEMA 4 Completo + Libroal436796Noch keine Bewertungen
- Mecanismos de AutocontrolDokument10 SeitenMecanismos de AutocontrolYakelin VillanuevaNoch keine Bewertungen
- sinapsisDokument11 SeitensinapsisCiber YaaxNoch keine Bewertungen
- Sinapsis QuìmicaDokument16 SeitenSinapsis QuìmicaFiorela Zarate100% (1)
- 3-Sistema Nervioso IDokument9 Seiten3-Sistema Nervioso IJavier FernandezNoch keine Bewertungen
- Neurotransmisión esencialDokument59 SeitenNeurotransmisión esencialNATALIA VALENTINA ALARCONNoch keine Bewertungen
- Tarea 3.1 - SinapsisDokument3 SeitenTarea 3.1 - SinapsisJassmin LariosNoch keine Bewertungen
- Comunicacion Neuronal SinapsisDokument11 SeitenComunicacion Neuronal Sinapsisvictor_cordova_66100% (1)
- Unidad 2Dokument48 SeitenUnidad 2NANCY CECILIA ORTEGA MARTINEZNoch keine Bewertungen
- Señalización CelularDokument13 SeitenSeñalización CelularAmadeo CabelloNoch keine Bewertungen
- Clasificación Estructural de La SinapsisDokument6 SeitenClasificación Estructural de La SinapsisRaz ZanNoch keine Bewertungen
- SinapsisDokument41 SeitenSinapsisNilda Cordova SanchezNoch keine Bewertungen
- Seminario 3 SinapsisDokument10 SeitenSeminario 3 SinapsisStefanny VillaltaNoch keine Bewertungen
- Tema 8-Bloque II-Comunicacion Neuronal SinapsisDokument12 SeitenTema 8-Bloque II-Comunicacion Neuronal SinapsisVictoria PerezNoch keine Bewertungen
- Diapositivas Seminario Comunicacion CelularDokument28 SeitenDiapositivas Seminario Comunicacion CelularAna Flavia Roncal RuizNoch keine Bewertungen
- Sinapsis PDFDokument11 SeitenSinapsis PDFMely BarriosNoch keine Bewertungen
- Síntesis NeurofisiologíaDokument175 SeitenSíntesis NeurofisiologíaMichell MolinaNoch keine Bewertungen
- NeurotransmisoresDokument72 SeitenNeurotransmisoresAyuwoki Hee heeNoch keine Bewertungen
- Neuro 2019 Seguna ParteDokument32 SeitenNeuro 2019 Seguna ParteKaren CuencaNoch keine Bewertungen
- Sinapsis (Completar en Clase Presencial) PDFDokument3 SeitenSinapsis (Completar en Clase Presencial) PDFEliangel lacleNoch keine Bewertungen
- Resumo Completo Fisio 2Dokument62 SeitenResumo Completo Fisio 2wandscheerdanielliNoch keine Bewertungen
- Solemnes Fisio GeneralDokument28 SeitenSolemnes Fisio GeneralGabriela VelásquezNoch keine Bewertungen
- Cuestionario S.N.C. Emperatriz IngaDokument9 SeitenCuestionario S.N.C. Emperatriz IngaEmperatrizIngaHidalgoNoch keine Bewertungen
- Histologia PracticasDokument10 SeitenHistologia PracticastunimhayreniNoch keine Bewertungen
- ComunicacionNeuronal2023 PDFDokument37 SeitenComunicacionNeuronal2023 PDFLuis DeviaNoch keine Bewertungen
- Fisiología GeneralDokument442 SeitenFisiología GeneralGabriela VelásquezNoch keine Bewertungen
- TEMA 7 Sinapsis PDFDokument62 SeitenTEMA 7 Sinapsis PDFJerónimoNoch keine Bewertungen
- clase 3 la sinapsisDokument59 Seitenclase 3 la sinapsisValentina HurtadoNoch keine Bewertungen
- Sinapsis: Comunicación Interneuronal PDFDokument4 SeitenSinapsis: Comunicación Interneuronal PDFSofi GómezNoch keine Bewertungen
- NeurotransmisorDokument7 SeitenNeurotransmisorzapatacamila952Noch keine Bewertungen
- Resumen Neuro 2Dokument24 SeitenResumen Neuro 2Constanza BoveraNoch keine Bewertungen
- Resumen Neurociencia Tema 3Dokument16 SeitenResumen Neurociencia Tema 3Nayara Solano JiménezNoch keine Bewertungen
- Guias ResueltasDokument4 SeitenGuias ResueltasAliciaEstelaDemariaBaguNoch keine Bewertungen
- Comunicación NeuronalDokument11 SeitenComunicación NeuronalAntonella MontielNoch keine Bewertungen
- Sistema Endocrino - p1 FibDokument3 SeitenSistema Endocrino - p1 FibCIKS JuicesNoch keine Bewertungen
- La Sinapsis en El Sistema Nervioso CentralDokument6 SeitenLa Sinapsis en El Sistema Nervioso Centralfrancisco bautista perezNoch keine Bewertungen
- Sinapsis ResumenDokument9 SeitenSinapsis ResumenAniretac Neko Seyer50% (4)
- Apuntes Tema 6Dokument17 SeitenApuntes Tema 6vanessa.delgado.psicologiaNoch keine Bewertungen
- SinapsisDokument4 SeitenSinapsisCesar Duque50% (2)
- Clase 4. Biologia de La ConductaDokument14 SeitenClase 4. Biologia de La ConductaLisi GarciaNoch keine Bewertungen
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Von EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017Noch keine Bewertungen
- Aprenda Farmacología En Una SemanaVon EverandAprenda Farmacología En Una SemanaBewertung: 2 von 5 Sternen2/5 (1)
- Los Seres Vivos Dependen De La Mecánica CuánticaVon EverandLos Seres Vivos Dependen De La Mecánica CuánticaNoch keine Bewertungen
- Normas Apa 6 EdiciónDokument14 SeitenNormas Apa 6 EdiciónLeidy CastrillónNoch keine Bewertungen
- Yo Antes y Después de La JubilaciónDokument1 SeiteYo Antes y Después de La JubilaciónDiego AmayaNoch keine Bewertungen
- Guia de EstudioDokument7 SeitenGuia de EstudioDiego AmayaNoch keine Bewertungen
- Ser CreativoDokument22 SeitenSer CreativoDiego AmayaNoch keine Bewertungen
- 10el Sujeto CognoscenteDokument17 Seiten10el Sujeto CognoscenteDiego AmayaNoch keine Bewertungen
- CirculoDokument23 SeitenCirculoDiego Amaya100% (1)
- Lenguaje y ComunicaciónDokument41 SeitenLenguaje y ComunicaciónDiego AmayaNoch keine Bewertungen
- Esquemas Modelo RenasciDokument2 SeitenEsquemas Modelo RenasciDiego AmayaNoch keine Bewertungen
- Test Gestáltico Visomotor de Lauretta BenderDokument10 SeitenTest Gestáltico Visomotor de Lauretta BenderDiego AmayaNoch keine Bewertungen
- Comienzo Del ConductismoDokument12 SeitenComienzo Del ConductismoDiego AmayaNoch keine Bewertungen
- Ser CreativoDokument22 SeitenSer CreativoDiego AmayaNoch keine Bewertungen
- Dibujo de La Figura Humana de Karen MachoverDokument4 SeitenDibujo de La Figura Humana de Karen MachoverDiego AmayaNoch keine Bewertungen
- Introduccion Al ConductismoDokument8 SeitenIntroduccion Al ConductismoDiego AmayaNoch keine Bewertungen
- Registros Sobre La Modificación de La ConductaDokument13 SeitenRegistros Sobre La Modificación de La ConductaDiego AmayaNoch keine Bewertungen
- 02 Trastornos Del AdultoDokument24 Seiten02 Trastornos Del AdultoDiego AmayaNoch keine Bewertungen
- Bases teóricas pruebas proyectivasDokument10 SeitenBases teóricas pruebas proyectivasDiego AmayaNoch keine Bewertungen
- Dibujo de La FamiliaDokument5 SeitenDibujo de La FamiliaDiego AmayaNoch keine Bewertungen
- La Autobiografía Como Técnica ProyectivaDokument7 SeitenLa Autobiografía Como Técnica ProyectivaDiego AmayaNoch keine Bewertungen
- Flores de BachDokument10 SeitenFlores de BachDiego AmayaNoch keine Bewertungen
- Bases Teóricas para El PsicodiagnósticoDokument11 SeitenBases Teóricas para El PsicodiagnósticoDiego AmayaNoch keine Bewertungen
- Bases teóricas pruebas proyectivasDokument10 SeitenBases teóricas pruebas proyectivasDiego AmayaNoch keine Bewertungen
- Trastorno Reactivo de La Vinculación en La InfanciaDokument14 SeitenTrastorno Reactivo de La Vinculación en La InfanciaDiego AmayaNoch keine Bewertungen
- La Teoría Centrada en La Persona de RogersDokument38 SeitenLa Teoría Centrada en La Persona de RogersDiego AmayaNoch keine Bewertungen
- 01 Trastornos de La InfanciaDokument35 Seiten01 Trastornos de La InfanciaDiego AmayaNoch keine Bewertungen
- Bases Teóricas para El PsicodiagnósticoDokument11 SeitenBases Teóricas para El PsicodiagnósticoDiego AmayaNoch keine Bewertungen
- Cat-A y Cat-H InfantilDokument39 SeitenCat-A y Cat-H InfantilDiego Amaya100% (8)
- Tipos Diagnósitco Factores de Riesgo 10 20 30 40 50 Señales de Alarma 10 20 30 40 50 TratamientoDokument56 SeitenTipos Diagnósitco Factores de Riesgo 10 20 30 40 50 Señales de Alarma 10 20 30 40 50 TratamientoDiego AmayaNoch keine Bewertungen
- Flores de BachDokument10 SeitenFlores de BachDiego AmayaNoch keine Bewertungen
- Otro Jueves CobardeDokument2 SeitenOtro Jueves CobardeDiego Amaya100% (1)
- Nutricion en SepsisDokument34 SeitenNutricion en SepsisChristian Javier Tana BasantesNoch keine Bewertungen
- Funcion de Nutricion en Seres VivosDokument17 SeitenFuncion de Nutricion en Seres VivosLuz GómezNoch keine Bewertungen
- Cardiología Precarga y PotcargaDokument3 SeitenCardiología Precarga y PotcargaDaniel ValenzuelaNoch keine Bewertungen
- Carne y Productos Cárnicos Como Fuente de PéptidosDokument10 SeitenCarne y Productos Cárnicos Como Fuente de PéptidosJoséVictorFlorezRosarioNoch keine Bewertungen
- Exposicion de Reflexlogia!!!Dokument25 SeitenExposicion de Reflexlogia!!!Liliana Andrea Villamarin LopezNoch keine Bewertungen
- Arrugas de La PielDokument5 SeitenArrugas de La PielHugo Villca LutinoNoch keine Bewertungen
- GRUPO 43 Tarea 4 - Metabolismo Grupo - 43Dokument26 SeitenGRUPO 43 Tarea 4 - Metabolismo Grupo - 43yeris selena mora marroquin33% (3)
- Apuntes Sistema RespiratorioDokument33 SeitenApuntes Sistema Respiratoriomaria de los angeles sanchez ortegaNoch keine Bewertungen
- Principales Inmovilizaciones de Yeso. Ortesis y ProtesisDokument42 SeitenPrincipales Inmovilizaciones de Yeso. Ortesis y Protesisalmaomiguel12Noch keine Bewertungen
- Arcos Faringeos Resumen SomaDokument4 SeitenArcos Faringeos Resumen SomaARACELY AMARONoch keine Bewertungen
- 2020 Neurodegeneracion Cerebro ContaminacionDokument207 Seiten2020 Neurodegeneracion Cerebro ContaminacionlidysNoch keine Bewertungen
- Stress Relief ComplexDokument11 SeitenStress Relief ComplexshakleeprNoch keine Bewertungen
- Ictericia neonatal: causas, diagnóstico y tratamientoDokument21 SeitenIctericia neonatal: causas, diagnóstico y tratamientoMario MagueyalNoch keine Bewertungen
- Biomecánica de CaderaDokument23 SeitenBiomecánica de CaderaMary CamachoNoch keine Bewertungen
- Vitamina B4 o AdeninaDokument6 SeitenVitamina B4 o AdeninaAnonymous ybCzeSJ7CNoch keine Bewertungen
- Body Piercing Profesional 22 27961 CompletoDokument40 SeitenBody Piercing Profesional 22 27961 CompletoLorraine ManriqueNoch keine Bewertungen
- Ejercicios de BHEDokument10 SeitenEjercicios de BHENoelia Lisbeth Castillo RuizNoch keine Bewertungen
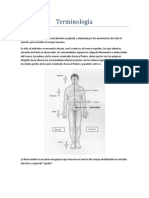
- Terminología Educación para La SaludDokument8 SeitenTerminología Educación para La SaludNatalia Sánchez100% (2)
- Labio Leporino y Paladar HendidoDokument11 SeitenLabio Leporino y Paladar HendidoLennyn Galindo RojasNoch keine Bewertungen
- Transición Recién NacidoDokument9 SeitenTransición Recién NacidoAnBCNoch keine Bewertungen
- Averts PDFDokument1 SeiteAverts PDFAlejandro RamirezNoch keine Bewertungen
- 2003 n1 Revision Regulacion Del Apetito Nuevos ConceptosDokument8 Seiten2003 n1 Revision Regulacion Del Apetito Nuevos ConceptosFederico TedescoNoch keine Bewertungen
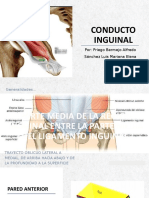
- Conducto inguinal anatómicoDokument13 SeitenConducto inguinal anatómicoMariana Elena SánchezNoch keine Bewertungen
- HIPERTENSION ARTERiALDokument5 SeitenHIPERTENSION ARTERiALErickson Zeña Paiva100% (1)
- Cavidad Abdominal y Sistema Digestivo InfradiafragmáticoDokument10 SeitenCavidad Abdominal y Sistema Digestivo InfradiafragmáticoEmily SeijasNoch keine Bewertungen
- Pae CarmenDokument26 SeitenPae CarmenMaguita de OzNoch keine Bewertungen
- Heredado: Condiciones Asociadas Con Un Gran Número de AcantocitosDokument25 SeitenHeredado: Condiciones Asociadas Con Un Gran Número de AcantocitosjoseNoch keine Bewertungen
- Músculos de La MimicaDokument3 SeitenMúsculos de La MimicaSamia Ruiz100% (1)
- Características de Los Seres VivosDokument3 SeitenCaracterísticas de Los Seres VivosAristoteles Socrates Moreno GoñeNoch keine Bewertungen
- Medrano Jimenez Jaime Abraham 2019081000741 PDFDokument4 SeitenMedrano Jimenez Jaime Abraham 2019081000741 PDFMaria AcevesNoch keine Bewertungen