Das könnte Ihnen auch gefallen
- Ebook Gratuito Receitas Ganhar Massa MuscularDokument16 SeitenEbook Gratuito Receitas Ganhar Massa MuscularMax Oliveira100% (3)
- Estrutura Celular RESUMODokument2 SeitenEstrutura Celular RESUMOMariana RodriguesNoch keine Bewertungen
- Poder simbólico e dominação de classesDokument3 SeitenPoder simbólico e dominação de classesAndrey ViegasNoch keine Bewertungen
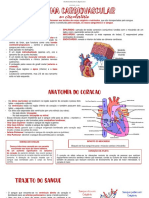
- Sistema CardiovascularDokument4 SeitenSistema CardiovascularEloiza CecíliaNoch keine Bewertungen
- Gimnospermas na CaatingaDokument24 SeitenGimnospermas na CaatingaOxe Biologia100% (1)
- HemipteraDokument31 SeitenHemipteraJessika Bárbara Lima100% (1)
- Organelas e estruturas celulares eucariontesDokument28 SeitenOrganelas e estruturas celulares eucariontesDouglas NobreNoch keine Bewertungen
- BCM-unidade III 1-2Dokument86 SeitenBCM-unidade III 1-2Edmond TNoch keine Bewertungen
- Resumão de BiologiaDokument13 SeitenResumão de BiologiajanaliahNoch keine Bewertungen
- Membrana celular 2024Dokument43 SeitenMembrana celular 2024azaelNoch keine Bewertungen
- Organização CelularDokument28 SeitenOrganização CelularEric Fernandes100% (2)
- Células eucariontes: estrutura e funçãoDokument5 SeitenCélulas eucariontes: estrutura e funçãoLucas NevesNoch keine Bewertungen
- Membrana BiologicaDokument5 SeitenMembrana BiologicaMaria Eduarda Padilha BaltazarNoch keine Bewertungen
- CitologiaDokument8 SeitenCitologiaericc-13hotmail.comNoch keine Bewertungen
- MI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaDokument4 SeitenMI 1º ANO Unid 3 - Célula - Cap. 7a - Membrana PlasmáticaEriczinNoch keine Bewertungen
- 1 - Organização Estrutural e Funcional Das CélulasDokument17 Seiten1 - Organização Estrutural e Funcional Das CélulasEduardo MatosNoch keine Bewertungen
- Resumo - GenéticaDokument16 SeitenResumo - Genéticaisabeljulia968Noch keine Bewertungen
- 3 Mpa Citologia Membrana Plasmática Profa. Marlene BoccattoDokument51 Seiten3 Mpa Citologia Membrana Plasmática Profa. Marlene BoccattoÉrin MartinsNoch keine Bewertungen
- Aula 3 Superficie Celular Organizacao Molecular e Funcao 2022Dokument49 SeitenAula 3 Superficie Celular Organizacao Molecular e Funcao 2022Emerson FranciscoNoch keine Bewertungen
- Membrana celular e permeabilidadeDokument33 SeitenMembrana celular e permeabilidadehendrya08Noch keine Bewertungen
- Fund. Biol. - Aula 4 - Membrana CelularDokument22 SeitenFund. Biol. - Aula 4 - Membrana CelularizabellemvalenteNoch keine Bewertungen
- CURSO DE EDUCAÇÃO FÍSICADokument6 SeitenCURSO DE EDUCAÇÃO FÍSICAGrazi MoreiraNoch keine Bewertungen
- NST-biologia-Citologia ZeroDokument13 SeitenNST-biologia-Citologia ZeroGabriel cardosooNoch keine Bewertungen
- Revisao para Prova de CitologiaDokument30 SeitenRevisao para Prova de CitologiaAdilson Marcos de SouzaNoch keine Bewertungen
- Bio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaDokument7 SeitenBio Aula Célula - Transporte - Divisão Celular - Embriologia - HistologiaMatheus PadilhaNoch keine Bewertungen
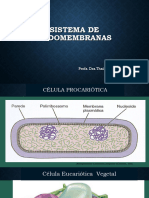
- Sistema de EndomembranasDokument26 SeitenSistema de Endomembranascarmita geronimo garciaNoch keine Bewertungen
- Biologia Celular IntroduçãoDokument38 SeitenBiologia Celular IntroduçãoMaxwell M. RibeiroNoch keine Bewertungen
- Material CITOLOGIA ORGANELASDokument10 SeitenMaterial CITOLOGIA ORGANELASScaryvanNoch keine Bewertungen
- Citologia e Transportes de MembranasDokument67 SeitenCitologia e Transportes de MembranasWalkyria Rodrigues RamosNoch keine Bewertungen
- Membrana Plasmática: Estrutura, Funções e TransporteDokument5 SeitenMembrana Plasmática: Estrutura, Funções e TransporteMoises FerroNoch keine Bewertungen
- Apostila de Biologia - Vol 2Dokument38 SeitenApostila de Biologia - Vol 2Biologia Concurso Vestibular100% (2)
- Transporte de substâncias nas célulasDokument18 SeitenTransporte de substâncias nas célulasMárcia JeteioNoch keine Bewertungen
- Exercicio Citologia RespondidoDokument10 SeitenExercicio Citologia RespondidoWanessa TeófilaNoch keine Bewertungen
- Modulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Dokument13 SeitenModulo 2 Introducao A Citologia e Membrana Plasmatica - Doc - 20240314 - 113055 - 0000Akiane FelixNoch keine Bewertungen
- Membrana celular: funções e estruturaDokument20 SeitenMembrana celular: funções e estruturaMario Maganizo JuniorNoch keine Bewertungen
- Componentes e estruturas do núcleo celularDokument7 SeitenComponentes e estruturas do núcleo celularohohnãoprecisasdesabermeunomeNoch keine Bewertungen
- Membrana celular permeabilidadeDokument21 SeitenMembrana celular permeabilidadeAriadne SiqueiraNoch keine Bewertungen
- Estudo DirigidoDokument4 SeitenEstudo DirigidoJoao Pedro Soares Amaral SousaNoch keine Bewertungen
- Bases Da Biologia Celular e Envoltórios CelularesDokument53 SeitenBases Da Biologia Celular e Envoltórios Celularesaline.fernandes11Noch keine Bewertungen
- Membrana Celular: Características e FunçõesDokument24 SeitenMembrana Celular: Características e FunçõesAna Barth100% (1)
- Biocel - PHDokument70 SeitenBiocel - PHpaulohnunesNoch keine Bewertungen
- Sist - ENDOMEMBRANA 2023 - CBiomedDokument46 SeitenSist - ENDOMEMBRANA 2023 - CBiomedavenidadepirocaNoch keine Bewertungen
- CitologiaDokument45 SeitenCitologiaNikilitosNoch keine Bewertungen
- Citologia QuestionarioDokument6 SeitenCitologia Questionariogiovanna siqueiraNoch keine Bewertungen
- Resumo - Unidade I BHEDokument5 SeitenResumo - Unidade I BHEAmanda GonçalvesNoch keine Bewertungen
- Obtencaomateriamembrana 202122Dokument30 SeitenObtencaomateriamembrana 202122Ana FernandesNoch keine Bewertungen
- Trabalaho Membrana PlasmáticaDokument16 SeitenTrabalaho Membrana PlasmáticaRonny GoisNoch keine Bewertungen
- Organização Molecular da Membrana CelularDokument36 SeitenOrganização Molecular da Membrana CelularIsabela Honorato50% (2)
- UN3Dokument13 SeitenUN3Gildo Flório JúniorNoch keine Bewertungen
- Membrana Plasmática. Digestão IntracelularDokument27 SeitenMembrana Plasmática. Digestão IntracelularBruno PereiraNoch keine Bewertungen
- Introdução Ao Estudo Da CélulaDokument25 SeitenIntrodução Ao Estudo Da Céluladrifrancas88100% (10)
- Células eucariontes: estrutura e componentes da membrana plasmáticaDokument18 SeitenCélulas eucariontes: estrutura e componentes da membrana plasmáticaLucas NevesNoch keine Bewertungen
- O conceito e componentes da célulaDokument8 SeitenO conceito e componentes da célulaRoberto Eduardo VieiraNoch keine Bewertungen
- CITOLOGIA Classificação Das CelulasDokument53 SeitenCITOLOGIA Classificação Das Celulascamila.alvesNoch keine Bewertungen
- 4º Relatório de Biologia CelularDokument11 Seiten4º Relatório de Biologia CelularalinemsantanaNoch keine Bewertungen
- MEMBRANA CELULAR Aula 3Dokument44 SeitenMEMBRANA CELULAR Aula 3Isaura ChiyeckNoch keine Bewertungen
- Membrana Plasmatica BioquimDokument83 SeitenMembrana Plasmatica Bioquimflavia douraoNoch keine Bewertungen
- Questionário I - Biologia CelularDokument6 SeitenQuestionário I - Biologia CelularBianca Sa100% (6)
- Constituintes CelularesDokument36 SeitenConstituintes CelularesHeitor RamosNoch keine Bewertungen
- Membrana Plasmática e Transporte Pela MembranaDokument63 SeitenMembrana Plasmática e Transporte Pela MembranaVitor MarinhoNoch keine Bewertungen
- Aula de RevisãoDokument36 SeitenAula de RevisãoHéllem SimoneNoch keine Bewertungen
- Sistema de endomembranas: componentes y funcionesDokument9 SeitenSistema de endomembranas: componentes y funcionesEdgardo Andrés Bilbao MuñozNoch keine Bewertungen
- Mastectomia Radical Modificada e Tecnicas de Cirugia ConservadoraDokument1 SeiteMastectomia Radical Modificada e Tecnicas de Cirugia ConservadoraAndrey ViegasNoch keine Bewertungen
- Relações entre postura corporal e sistema estomatognáticoDokument10 SeitenRelações entre postura corporal e sistema estomatognáticoAndrey ViegasNoch keine Bewertungen
- Regulamento Geral OABDokument4 SeitenRegulamento Geral OABAndrey ViegasNoch keine Bewertungen
- Cap 17 ReferenciaDokument46 SeitenCap 17 ReferenciaAndrey ViegasNoch keine Bewertungen
- Biof - Sica Da Vis - oDokument54 SeitenBiof - Sica Da Vis - oAndrey ViegasNoch keine Bewertungen
- DietaaaDokument1 SeiteDietaaaAndrey ViegasNoch keine Bewertungen
- Ganhe Trabalhando CasaDokument9 SeitenGanhe Trabalhando CasaAndrey Viegas0% (1)
- Aula 32 - Lubrificação Industrial IIDokument9 SeitenAula 32 - Lubrificação Industrial IIDouglas Gomes100% (12)
- Manutenção PreditivaDokument5 SeitenManutenção PreditivaAndrey ViegasNoch keine Bewertungen
- ACC1 - Prova de Teste 7° Ano 1BIMDokument4 SeitenACC1 - Prova de Teste 7° Ano 1BIMJoão Paulo GaldinoNoch keine Bewertungen
- Avaliação do projeto RPG na Biblioteca MunicipalDokument11 SeitenAvaliação do projeto RPG na Biblioteca MunicipalThiago Rodrigues Nascimento100% (1)
- Entre Índios AruansDokument98 SeitenEntre Índios AruansAna LatgeNoch keine Bewertungen
- Sistema Tegumentar: Funções e Estrutura da PeleDokument39 SeitenSistema Tegumentar: Funções e Estrutura da PeleMaria Stella FernandesNoch keine Bewertungen
- Pornografia e SexualidadeDokument24 SeitenPornografia e SexualidadeRoneldo Moura da SilvaNoch keine Bewertungen
- Design EmocionalDokument10 SeitenDesign EmocionalPedroNoch keine Bewertungen
- Carr & Shepherd 2006.en - PTDokument10 SeitenCarr & Shepherd 2006.en - PTBrenaOliveiraNoch keine Bewertungen
- Pense Menos, viva mais - Nick TrentonDokument182 SeitenPense Menos, viva mais - Nick TrentonBartolomeu SantosNoch keine Bewertungen
- Resumos Temperatura No Crescimento Microbiano e Metabolismo MicrobianoDokument59 SeitenResumos Temperatura No Crescimento Microbiano e Metabolismo MicrobianoDiogo CorreiaNoch keine Bewertungen
- Desenvolvimento Cognitivo - Jean PiagetDokument11 SeitenDesenvolvimento Cognitivo - Jean PiagetJosé Hiroshi Taniguti100% (24)
- O Tempo GeológicoDokument4 SeitenO Tempo GeológicoRoger LimaNoch keine Bewertungen
- Dor neuropática: mecanismos e classificaçãoDokument9 SeitenDor neuropática: mecanismos e classificaçãoPedro Augusto Arariba NevesNoch keine Bewertungen
- Sumidouro e Sifão Anti OdorDokument2 SeitenSumidouro e Sifão Anti Odorrita_mendes_1Noch keine Bewertungen
- Aula Disturbios Acido BasicoDokument26 SeitenAula Disturbios Acido BasicoCamila GarciaNoch keine Bewertungen
- Os Manguezais Nos Livros de Ciências NaturaisDokument61 SeitenOs Manguezais Nos Livros de Ciências NaturaisJose FinaNoch keine Bewertungen
- Teoria Do Autocuidado de OremDokument7 SeitenTeoria Do Autocuidado de OremAlbertina SousaNoch keine Bewertungen
- Fundbio Aula3Dokument10 SeitenFundbio Aula3Jose Barbosa Da Silva FilhoNoch keine Bewertungen
- Caça Ao Erro OrtograficoDokument2 SeitenCaça Ao Erro OrtograficoEduardoNoch keine Bewertungen
- João Antônio - Corpo-A-Corpo Com A VidaDokument6 SeitenJoão Antônio - Corpo-A-Corpo Com A Vidaakupac100% (1)
- Trombose Venosa Profunda: Deep Venous ThrombosisDokument7 SeitenTrombose Venosa Profunda: Deep Venous ThrombosisMedPBNoch keine Bewertungen
- Bahiana - Área de Saúde - Prova - 2021 - 2Dokument18 SeitenBahiana - Área de Saúde - Prova - 2021 - 2Bruna SabaNoch keine Bewertungen
- Doença crítica crônica: salvando vidas ou criando vítimasDokument9 SeitenDoença crítica crônica: salvando vidas ou criando vítimasKarol VersianiNoch keine Bewertungen
- Aplicabilidade Da Pimenta (Capsaicina) Na EstéticaDokument11 SeitenAplicabilidade Da Pimenta (Capsaicina) Na EstéticamjmorcelliNoch keine Bewertungen
- Alga Marrom (Ascophyllum Nodosum) para Alevinos de Tilápia Do NiloDokument8 SeitenAlga Marrom (Ascophyllum Nodosum) para Alevinos de Tilápia Do NiloJosué ItaloNoch keine Bewertungen
- Técnica de Esterilização Térmica de AlimentosDokument6 SeitenTécnica de Esterilização Térmica de AlimentosThaissa Ilana LimaNoch keine Bewertungen
- Planner Resumov ENEMDokument8 SeitenPlanner Resumov ENEMJulia da Silva NogueiraNoch keine Bewertungen
- Tudo sobre AromaterapiaDokument31 SeitenTudo sobre AromaterapiaAdao BentoNoch keine Bewertungen