Das könnte Ihnen auch gefallen
- Proteínas GlobularesDokument17 SeitenProteínas GlobularesArturo Sandoval ahuanariNoch keine Bewertungen
- Grupo HemoDokument20 SeitenGrupo HemoDavid RecharteNoch keine Bewertungen
- Guia de Estudio y Autorregulacion BiomoleculasDokument30 SeitenGuia de Estudio y Autorregulacion BiomoleculasFelipe Antonio Urra DonosoNoch keine Bewertungen
- Los Elementos de La Vida Todos LosDokument3 SeitenLos Elementos de La Vida Todos LosmalditonNoch keine Bewertungen
- ProteínasDokument5 SeitenProteínasLizeth Monserrat RomeroNoch keine Bewertungen
- Las porfirinas: estructura, composición y aplicacionesDokument55 SeitenLas porfirinas: estructura, composición y aplicacionesMarín GENoch keine Bewertungen
- Proteínas y EnzimasDokument5 SeitenProteínas y EnzimasLizeth Monserrat RomeroNoch keine Bewertungen
- Bloque Ii Biologia I 2023Dokument39 SeitenBloque Ii Biologia I 2023Yeshua C MuñozNoch keine Bewertungen
- Temas Selectos de QuímicaDokument25 SeitenTemas Selectos de QuímicaScl BernabeNoch keine Bewertungen
- 2 BioelementosDokument5 Seiten2 BioelementosTienda OkkusNoch keine Bewertungen
- Los Elementos de La VidaDokument8 SeitenLos Elementos de La VidaSilvio MarianiNoch keine Bewertungen
- Sintesis Del Grupo Hemo 1Dokument4 SeitenSintesis Del Grupo Hemo 1tomasNoch keine Bewertungen
- Bioelementos MIADokument18 SeitenBioelementos MIACEBA Raúl Paredes EspinozaNoch keine Bewertungen
- Citocromo P450Dokument34 SeitenCitocromo P450Miguel Angel Gómez Ramos100% (1)
- Importancia Biologica de Las ProteinasDokument9 SeitenImportancia Biologica de Las Proteinasjuan angelNoch keine Bewertungen
- Reconocimiento de BioelementosDokument12 SeitenReconocimiento de BioelementosAlejandra Garcia ObregonNoch keine Bewertungen
- Procesos BiológicosDokument24 SeitenProcesos BiológicosARELI JOCABETH VALLADARES CABRALNoch keine Bewertungen
- Bioquimica Tarea 07 SepDokument2 SeitenBioquimica Tarea 07 SepLAURA ISABELLA CÁRDENAS GUARNIZONoch keine Bewertungen
- BIOLOGÍADokument11 SeitenBIOLOGÍANueva CuentaNoch keine Bewertungen
- Guía Nº2 Biomoleculas, Bioelementos, Vitaminas Bioquimica Teoria PDFDokument7 SeitenGuía Nº2 Biomoleculas, Bioelementos, Vitaminas Bioquimica Teoria PDFCAROL JOHANA MONTA�EZ JAIMES100% (1)
- Biomoleculas y Su Interacción en La CélulaDokument53 SeitenBiomoleculas y Su Interacción en La CélulaIvan MontañoNoch keine Bewertungen
- Proteínas: estructura y funcionesDokument5 SeitenProteínas: estructura y funcionesLeoNoch keine Bewertungen
- Bioelementos esenciales para la vida: Carbono, Oxígeno, Nitrógeno y másDokument3 SeitenBioelementos esenciales para la vida: Carbono, Oxígeno, Nitrógeno y másValery MerchánNoch keine Bewertungen
- Citocromo P450Dokument5 SeitenCitocromo P450Cindy SalazarNoch keine Bewertungen
- Componentes de la sangreDokument13 SeitenComponentes de la sangreNacho SanchezNoch keine Bewertungen
- Bloque 3 Cuaderno de ActividadesDokument21 SeitenBloque 3 Cuaderno de ActividadesXtiianNoch keine Bewertungen
- Biotransformacion Toxicos Orozco Morales Lizbeth Gpe.Dokument14 SeitenBiotransformacion Toxicos Orozco Morales Lizbeth Gpe.Angel Rxn0% (1)
- Síntesis Del HemoDokument8 SeitenSíntesis Del HemoHenryk BazalarNoch keine Bewertungen
- Biolementos PrimariosDokument11 SeitenBiolementos PrimariosLifeC4TNoch keine Bewertungen
- Introduccion A La Quimica de Los AlimentDokument11 SeitenIntroduccion A La Quimica de Los Alimentluis murielNoch keine Bewertungen
- Importancia del nitrógeno en las proteínas y seres vivosDokument6 SeitenImportancia del nitrógeno en las proteínas y seres vivosSUANSEDNoch keine Bewertungen
- Base Quimica de La Vida IntroduccionDokument28 SeitenBase Quimica de La Vida IntroduccionAlba TorresNoch keine Bewertungen
- Aminoácidos y ProteinasDokument6 SeitenAminoácidos y ProteinasAsami AkitaNoch keine Bewertungen
- Guia 1 Bioelementos y BiocompuestosDokument7 SeitenGuia 1 Bioelementos y BiocompuestosGanjajuank GanjajuankNoch keine Bewertungen
- 01 Lib Bioquimica 6Dokument6 Seiten01 Lib Bioquimica 6Andrea Cubero MontesNoch keine Bewertungen
- GuzmanAguilar FelipeGuillermo M14S3AI6Dokument4 SeitenGuzmanAguilar FelipeGuillermo M14S3AI6felipe guillermo guzman aguilar50% (2)
- Moléculas metabólicasDokument13 SeitenMoléculas metabólicasClaudia Casanova MuñozNoch keine Bewertungen
- Guía de las biomoléculas: Características y funcionesDokument4 SeitenGuía de las biomoléculas: Características y funcionesOscar Yamid JovenNoch keine Bewertungen
- Porfirira Eritropoyética Congénita - Alejandro P. BojórquezDokument9 SeitenPorfirira Eritropoyética Congénita - Alejandro P. BojórquezAlejandro Pesqueira BojorquezNoch keine Bewertungen
- Unidad I AminoácidosDokument10 SeitenUnidad I AminoácidosRaul Eduardo Medori RomeroNoch keine Bewertungen
- Laboratorio PRACTICA BIOMOLECULASDokument9 SeitenLaboratorio PRACTICA BIOMOLECULASCM3576% (25)
- Clasificacion de Lo BioelementosDokument7 SeitenClasificacion de Lo BioelementoslizaNoch keine Bewertungen
- ProteinasDokument12 SeitenProteinasAlejandro LunaNoch keine Bewertungen
- BiologiaDokument17 SeitenBiologiaanais gacituaNoch keine Bewertungen
- Bioelementos y Grupos FuncionalesDokument3 SeitenBioelementos y Grupos FuncionalesJohnny Fer ÆNoch keine Bewertungen
- Bases Moleculares y Celulares de La VidaDokument15 SeitenBases Moleculares y Celulares de La VidajheniferNoch keine Bewertungen
- Vitaminas y Oxidorreductasas AntioxidantesDokument30 SeitenVitaminas y Oxidorreductasas AntioxidantesApril CollinsNoch keine Bewertungen
- Bioquímica clínica: metabolismo y combustiblesDokument118 SeitenBioquímica clínica: metabolismo y combustiblesXanny Espinoza CastroNoch keine Bewertungen
- Cómo Estamos Formados Los Seres HumanosDokument2 SeitenCómo Estamos Formados Los Seres HumanosHarimander KaurNoch keine Bewertungen
- Composición química de los seres vivosDokument66 SeitenComposición química de los seres vivosLuis Choquecota MendozaNoch keine Bewertungen
- Cloro y ProteínasDokument3 SeitenCloro y ProteínasIssa BelNoch keine Bewertungen
- Biomoléculas ExamenDokument65 SeitenBiomoléculas ExamenNERYS ESTELLA RAMOS BOHORQUEZNoch keine Bewertungen
- Bio. (01) Bioelementos y Agua 1 - 10Dokument10 SeitenBio. (01) Bioelementos y Agua 1 - 10ALEXANDER DAVID LOPEZ ESCOBEDONoch keine Bewertungen
- Que Es Una Biomolécula ExposicionDokument4 SeitenQue Es Una Biomolécula ExposicionMARIA USTARIZ FRAGOZONoch keine Bewertungen
- Aminoacidos y PeptidosDokument34 SeitenAminoacidos y PeptidosLucas FlorencioNoch keine Bewertungen
- Biomoleculas ProteinasDokument42 SeitenBiomoleculas ProteinasIvan GonzalezNoch keine Bewertungen
- PuerperioDokument26 SeitenPuerperiogabriela silvaNoch keine Bewertungen
- Cancer de OvarioDokument10 SeitenCancer de Ovariogabriela silvaNoch keine Bewertungen
- Cáncer de CervixDokument19 SeitenCáncer de Cervixgabriela silvaNoch keine Bewertungen
- Coordinadores LogísticaDokument5 SeitenCoordinadores Logísticagabriela silvaNoch keine Bewertungen
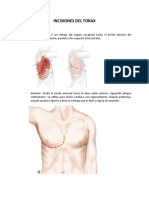
- Incisiones Del ToraxDokument4 SeitenIncisiones Del Toraxgabriela silvaNoch keine Bewertungen
- Bachelor DiplomaDokument1 SeiteBachelor Diplomagabriela silvaNoch keine Bewertungen
- Procesos en ToraxDokument5 SeitenProcesos en ToraxMP PCNoch keine Bewertungen
- Uso Del Ácido Tranexámico en La Hemorragia PospartoDokument6 SeitenUso Del Ácido Tranexámico en La Hemorragia Pospartogabriela silvaNoch keine Bewertungen
- TUMORES DE HIGADO Y PERIAMPULARES FinalDokument36 SeitenTUMORES DE HIGADO Y PERIAMPULARES Finalgabriela silvaNoch keine Bewertungen
- Tumores de Partes BlandasDokument8 SeitenTumores de Partes Blandasgabriela silvaNoch keine Bewertungen
- Ins. Respiratoria ExpoDokument2 SeitenIns. Respiratoria Expogabriela silvaNoch keine Bewertungen
- Patologías de Piel en Pacientes PediatricosDokument51 SeitenPatologías de Piel en Pacientes Pediatricosgabriela silvaNoch keine Bewertungen
- Doc to U: Plataforma para agendar citas médicas y gestionar historias clínicasDokument2 SeitenDoc to U: Plataforma para agendar citas médicas y gestionar historias clínicasgabriela silvaNoch keine Bewertungen
- Mesa Terapeutica DX TuberculosisDokument5 SeitenMesa Terapeutica DX Tuberculosisgabriela silvaNoch keine Bewertungen
- Efecto de La Insulina Sobre El Metabolismo de Las Proteínas y El CrecimientoDokument5 SeitenEfecto de La Insulina Sobre El Metabolismo de Las Proteínas y El CrecimientoEmiiZambranoNoch keine Bewertungen
- Glosario BiologíaDokument2 SeitenGlosario BiologíaZeltzin CansecoNoch keine Bewertungen
- Sistemas K y V enzimas alostéricasDokument3 SeitenSistemas K y V enzimas alostéricasYeisyAlayo0% (1)
- Trabajo AminoacidosDokument38 SeitenTrabajo AminoacidosYojackson Pastran100% (1)
- Resumen GeneticaDokument165 SeitenResumen GeneticaFlorencia SierraNoch keine Bewertungen
- ApolipoproteínasDokument11 SeitenApolipoproteínasQuero AldíiNoch keine Bewertungen
- Cinetica EnzimaticaDokument3 SeitenCinetica EnzimaticaNIDIA JULISSA ALCALDE CARDENASNoch keine Bewertungen
- Trastornos de CoagulaciónDokument6 SeitenTrastornos de CoagulaciónValeria CarriónNoch keine Bewertungen
- Practica 2 AmilasaDokument3 SeitenPractica 2 AmilasaSara Yelitza Gonzalez RasgadoNoch keine Bewertungen
- Verificación de la actividad proteolítica de enzimas en zumos de frutas mediante gelatina como sustratoDokument5 SeitenVerificación de la actividad proteolítica de enzimas en zumos de frutas mediante gelatina como sustratoLeoncio Hilario UmiyauriNoch keine Bewertungen
- PlasmidosDokument23 SeitenPlasmidosBetiana BerriryNoch keine Bewertungen
- 6 Neurotransmisores y NeuromoduladoresDokument106 Seiten6 Neurotransmisores y NeuromoduladoresKarol Morales100% (1)
- Terapia Sanguínea Del Paciente - CoagulaciónDokument30 SeitenTerapia Sanguínea Del Paciente - CoagulaciónMonica CalderónNoch keine Bewertungen
- Biomoléculas OrgánicasDokument5 SeitenBiomoléculas OrgánicasEros 09Noch keine Bewertungen
- Citocromo C: transportador electrónico claveDokument10 SeitenCitocromo C: transportador electrónico claveItzel RiveroNoch keine Bewertungen
- DERMAPEN - Marzo 2022Dokument4 SeitenDERMAPEN - Marzo 2022Bárbara LettoNoch keine Bewertungen
- Monografia AminoacidosDokument11 SeitenMonografia AminoacidosLenny Alvarado Ponce75% (4)
- Bloque 2 BioquímicaDokument23 SeitenBloque 2 BioquímicaSergio SeseteNoch keine Bewertungen
- Enzimología PDFDokument28 SeitenEnzimología PDFBryan Esparza100% (1)
- Señalizacion Celular IiDokument40 SeitenSeñalizacion Celular IiFABIAN RODAS MENDOZANoch keine Bewertungen
- Cuestionario Bioquimica 4 de MayoDokument7 SeitenCuestionario Bioquimica 4 de MayoTiffany FongNoch keine Bewertungen
- Clase 9 - Metabolismo de La Glucosa - C Krebs - F OxidativaDokument31 SeitenClase 9 - Metabolismo de La Glucosa - C Krebs - F OxidativaConstanza Nicole Alarcon EcheverriaNoch keine Bewertungen
- Ruta Del ShikimatoDokument10 SeitenRuta Del ShikimatoAdrian Alberto López CenNoch keine Bewertungen
- Bioelementos y BiocompuestosDokument10 SeitenBioelementos y BiocompuestosZharick MantillaNoch keine Bewertungen
- Clase 5 - ProteinasDokument53 SeitenClase 5 - ProteinasDanton Miranda CabreraNoch keine Bewertungen
- Teoría Replicación y Transcripción Del ADN, Traducción Del ARN.Dokument37 SeitenTeoría Replicación y Transcripción Del ADN, Traducción Del ARN.Juan Torres GalindoNoch keine Bewertungen
- EnzimasDokument20 SeitenEnzimasEmi LareNoch keine Bewertungen
- Activación de AminoácidosDokument5 SeitenActivación de AminoácidosAaron VargasNoch keine Bewertungen
- Lipidos ComplejosDokument2 SeitenLipidos ComplejosGiselle Archaga MartinezNoch keine Bewertungen
- Transcripción Genética: ARN Polimerasa Sintetiza ARNDokument5 SeitenTranscripción Genética: ARN Polimerasa Sintetiza ARNkadhelNoch keine Bewertungen