Das könnte Ihnen auch gefallen
- Mapa Conceptual - MicrobiologiaDokument1 SeiteMapa Conceptual - MicrobiologiaKatherine Terrones0% (1)
- HERBARIODokument5 SeitenHERBARIOMaira Valentina TorresNoch keine Bewertungen
- T.1 A T.7 FarmacognosiaDokument112 SeitenT.1 A T.7 FarmacognosiaMaria PucholNoch keine Bewertungen
- 8 Farmacos AntiparasitariosDokument12 Seiten8 Farmacos AntiparasitariosJoseNoch keine Bewertungen
- Practica Equilibrio OsmoticoDokument13 SeitenPractica Equilibrio OsmoticoFer Mend EleNoch keine Bewertungen
- Hi Peru Rice MiaDokument23 SeitenHi Peru Rice MiaAleNoch keine Bewertungen
- Pam236 Plantas Medicinales Con QuinonasDokument6 SeitenPam236 Plantas Medicinales Con QuinonasLuiisa SánchezNoch keine Bewertungen
- GastrinaDokument2 SeitenGastrinaCarlos SorianoNoch keine Bewertungen
- Hoja de CocaDokument45 SeitenHoja de Cocaluz100% (2)
- Practica 1. Cinetica Del Etanol.Dokument14 SeitenPractica 1. Cinetica Del Etanol.Alvaro GarciaNoch keine Bewertungen
- Farmacognosia GB 12Dokument12 SeitenFarmacognosia GB 12Guillaume michiganNoch keine Bewertungen
- Antitusivos y MucolíticosDokument14 SeitenAntitusivos y MucolíticosMaría IsazaNoch keine Bewertungen
- Farmacologia Semana 2 Uw 2019Dokument41 SeitenFarmacologia Semana 2 Uw 2019Pedro Jacinto HerviasNoch keine Bewertungen
- Funcion Del ATPDokument15 SeitenFuncion Del ATPAlondra JimenezNoch keine Bewertungen
- Genetica BacterianaDokument44 SeitenGenetica Bacterianaanyita_0100% (3)
- Carbapenemes y MonobactamosDokument21 SeitenCarbapenemes y MonobactamosGabriela Prado GumielNoch keine Bewertungen
- Alcanos 2Dokument8 SeitenAlcanos 2Lili FuchsNoch keine Bewertungen
- TaxonomíaDokument5 SeitenTaxonomíaHouse VasquezNoch keine Bewertungen
- PENICILINASDokument28 SeitenPENICILINASManuel G. Oporta100% (1)
- Listado de Medicamentos PsicotropicosDokument5 SeitenListado de Medicamentos PsicotropicosMauricio De assis CanoNoch keine Bewertungen
- TEMA 4. - Introduccion A Los Comprimidos PDFDokument47 SeitenTEMA 4. - Introduccion A Los Comprimidos PDFLalos Serrano100% (1)
- Unidad 03 Organizacion Supramolecular y CelularDokument15 SeitenUnidad 03 Organizacion Supramolecular y CelularJavier GuerraNoch keine Bewertungen
- Efecto Diuretico de La OrtigaDokument7 SeitenEfecto Diuretico de La OrtigaPaolita TlvNoch keine Bewertungen
- Presentacion Chia.Dokument12 SeitenPresentacion Chia.Eliezer David Carrazco RodríguezNoch keine Bewertungen
- QUINOLONASDORS5Dokument36 SeitenQUINOLONASDORS5luisaNoch keine Bewertungen
- Clase 9Dokument20 SeitenClase 9Nexo CreativosNoch keine Bewertungen
- FarmacodinamicaDokument48 SeitenFarmacodinamicaLuis Eduardo Pilar SantosNoch keine Bewertungen
- Plantasmedicinalesyaromaticasmodulo1 150814032807 Lva1 App6891Dokument16 SeitenPlantasmedicinalesyaromaticasmodulo1 150814032807 Lva1 App6891Daniel MalpartidaNoch keine Bewertungen
- Excrecion Por BilisDokument2 SeitenExcrecion Por BilisJessica Giraldo CeledònNoch keine Bewertungen
- Metabolismo y Excreción DiaposDokument42 SeitenMetabolismo y Excreción DiaposStiward CamachoNoch keine Bewertungen
- Metabolismo de Los Antibióticos y XenobióticosDokument14 SeitenMetabolismo de Los Antibióticos y Xenobióticossantiago castiblancoNoch keine Bewertungen
- Toxicodinamica S3Dokument32 SeitenToxicodinamica S3Juan100% (1)
- La VacuolaDokument7 SeitenLa VacuolamaryajilaNoch keine Bewertungen
- Mapa Mental AbsorciónDokument1 SeiteMapa Mental AbsorciónErick RM100% (2)
- DIURETICOSDokument6 SeitenDIURETICOSAbigail M GomezNoch keine Bewertungen
- Mapas de Enzimas PancreaticasDokument2 SeitenMapas de Enzimas PancreaticasAngelShadow12Noch keine Bewertungen
- Farmacovigilancia y La Seguridad Del PacienteDokument44 SeitenFarmacovigilancia y La Seguridad Del PacientedavidNoch keine Bewertungen
- Macrolidos y TetraciclinasDokument25 SeitenMacrolidos y TetraciclinasJuanNoch keine Bewertungen
- 1 Historia de La Farmacia GlobalDokument32 Seiten1 Historia de La Farmacia GlobalMiranda BaezNoch keine Bewertungen
- Inflamación InmunoDokument11 SeitenInflamación Inmunosun_shine26Noch keine Bewertungen
- Esquema Ut 8 Formas FarmacéuticasDokument3 SeitenEsquema Ut 8 Formas FarmacéuticasVanesa Almenara NegrinNoch keine Bewertungen
- Definición y Origen de La FarmacologiaDokument13 SeitenDefinición y Origen de La FarmacologiaJennifer Salas CifuentesNoch keine Bewertungen
- SEMISOLIDOSDokument12 SeitenSEMISOLIDOSMarco Antonio Panta OlayaNoch keine Bewertungen
- La Farmacia ClínicaDokument4 SeitenLa Farmacia ClínicaAlfredo ApazaNoch keine Bewertungen
- Mapa Infraestructura de Laboratorios (Gavilanez)Dokument2 SeitenMapa Infraestructura de Laboratorios (Gavilanez)Sebásthian GavilanezNoch keine Bewertungen
- Trabajo Del Sida en WordDokument19 SeitenTrabajo Del Sida en WordIvan Martinez LopezNoch keine Bewertungen
- METABOLISMODokument31 SeitenMETABOLISMOMariana JiménezNoch keine Bewertungen
- Mapa Mental Enzimas Eimar Rivero 201O3Dokument1 SeiteMapa Mental Enzimas Eimar Rivero 201O3Eimar RiveroNoch keine Bewertungen
- FarmacosDokument18 SeitenFarmacosRayito ChavezNoch keine Bewertungen
- Músculos Del CráneoDokument4 SeitenMúsculos Del CráneoLuis Adolfo Ramirez AlonsoNoch keine Bewertungen
- Citocromo p450 2018-2Dokument29 SeitenCitocromo p450 2018-2PAULA NIETONoch keine Bewertungen
- RAMs 2018Dokument39 SeitenRAMs 2018GabrielaNoch keine Bewertungen
- UntitledDokument9 SeitenUntitledJesús Miguel Ramirez PinedoNoch keine Bewertungen
- TESIS LilianaDokument17 SeitenTESIS LilianaBioquimica UplaNoch keine Bewertungen
- Simpaticomimeticos y ColinergicosDokument83 SeitenSimpaticomimeticos y ColinergicosRocio Del Pilar Damian AlvaradoNoch keine Bewertungen
- Farmacologia IDokument106 SeitenFarmacologia IRaquel Ballenas100% (1)
- Ascaris LumbricoidesDokument1 SeiteAscaris LumbricoidesGabriel FloresNoch keine Bewertungen
- Paradigmas en MedicinaDokument29 SeitenParadigmas en MedicinaVíctor ArticaNoch keine Bewertungen
- 6 SaneamientoDokument2 Seiten6 SaneamientoKim Brayan O ANoch keine Bewertungen
- Clase 5 Canales IónicosDokument49 SeitenClase 5 Canales IónicosDiego Padilla SantosNoch keine Bewertungen
- Plan de Cuidados Paciente Quirurgico - 2010Dokument18 SeitenPlan de Cuidados Paciente Quirurgico - 2010Yeison Fernei Bastos SuaresNoch keine Bewertungen
- Guia para La Elaboración Informes de Servicio SocialDokument1 SeiteGuia para La Elaboración Informes de Servicio SocialJenifer CortésNoch keine Bewertungen
- Guia - ANEST.Hipertermia MalignaDokument5 SeitenGuia - ANEST.Hipertermia MalignaNancy Martinez VarelaNoch keine Bewertungen
- Franco 02 PDFDokument8 SeitenFranco 02 PDFchugodevilNoch keine Bewertungen
- 441GERDokument52 Seiten441GERJenifer CortésNoch keine Bewertungen
- NeumoniaDokument15 SeitenNeumoniaRositaRodriguezNoch keine Bewertungen
- Escala de GlasgowDokument2 SeitenEscala de GlasgowJenifer CortésNoch keine Bewertungen
- Farmacologia GeneralDokument214 SeitenFarmacologia GeneralCarolinalcmNoch keine Bewertungen
- GuillainBarrE R CENETEC Old2Dokument15 SeitenGuillainBarrE R CENETEC Old2alejandroNoch keine Bewertungen
- Unidad QX BDokument16 SeitenUnidad QX BJenifer CortésNoch keine Bewertungen
- Virginia Santos AlonsoDokument20 SeitenVirginia Santos AlonsoJenifer CortésNoch keine Bewertungen
- Depresion Escala HamiltonDokument4 SeitenDepresion Escala HamiltonLydia Ralfas Berges50% (2)
- Nom 005 Ssa2 1993Dokument78 SeitenNom 005 Ssa2 1993oziel gutierrezNoch keine Bewertungen
- Posiciones QXDokument1 SeitePosiciones QXJenifer CortésNoch keine Bewertungen
- 069 - Fisiología Del GustoDokument8 Seiten069 - Fisiología Del GustowilliamNoch keine Bewertungen
- Laringe y Faringe (1) AnatomiaDokument37 SeitenLaringe y Faringe (1) AnatomiaJenifer CortésNoch keine Bewertungen
- Lactancia MaternakhvDokument4 SeitenLactancia MaternakhvJenifer CortésNoch keine Bewertungen
- 9.1 Embarazo de Alto Riesgo MariaDokument1 Seite9.1 Embarazo de Alto Riesgo MariaJenifer CortésNoch keine Bewertungen
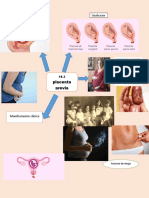
- Placenta PreviaDokument1 SeitePlacenta PreviaJenifer CortésNoch keine Bewertungen
- Mapa Conceptual KiuolDokument2 SeitenMapa Conceptual KiuolJenifer CortésNoch keine Bewertungen
- Diabetes PDFDokument16 SeitenDiabetes PDFMaria Gamboa AlajoNoch keine Bewertungen
- Clase 24Dokument16 SeitenClase 24Jesus Mendez RodriguezNoch keine Bewertungen
- Manual Enfermeria QuirurgicaDokument124 SeitenManual Enfermeria QuirurgicaJenifer Cortés100% (1)
- Manual de Procedimientos20012Dokument213 SeitenManual de Procedimientos20012Jenifer CortésNoch keine Bewertungen
- Revista Faso PDFDokument6 SeitenRevista Faso PDFAlenka MachadoNoch keine Bewertungen
- DigestivoDokument16 SeitenDigestivoAndrea GarcíaNoch keine Bewertungen
- Sistema RespiratorioDokument27 SeitenSistema RespiratorioitzelNoch keine Bewertungen
- TEMA 8a ProfesionDokument33 SeitenTEMA 8a ProfesionYisabel RondonNoch keine Bewertungen
- SistemaDokument21 SeitenSistemaLeonardo Andrs AveigaNoch keine Bewertungen
- ¿Cómo La Biotecnología Está Beneficiando A La Industria Alimentaria en Perú (2015-2020) ?Dokument7 Seiten¿Cómo La Biotecnología Está Beneficiando A La Industria Alimentaria en Perú (2015-2020) ?JOSE ALEJANDRO SAENZ PRINCIPENoch keine Bewertungen
- Lineas de InvestigaciónDokument7 SeitenLineas de InvestigaciónmgfidelNoch keine Bewertungen
- Guía Biología 9-1 Semana 12Dokument4 SeitenGuía Biología 9-1 Semana 12Edwins Chávez SarmientoNoch keine Bewertungen
- Semana 07 Terminado Expo BioDokument4 SeitenSemana 07 Terminado Expo BioSofiaNoch keine Bewertungen
- Proyecto Virus Equipo3Dokument8 SeitenProyecto Virus Equipo3Carlos Angel Arroyo QuiñonesNoch keine Bewertungen
- Bloque 1 Discu 5Dokument12 SeitenBloque 1 Discu 5Francisco GarcíaNoch keine Bewertungen
- D.G.L.Q - Laboratorio Sem14Dokument22 SeitenD.G.L.Q - Laboratorio Sem14Karol Astorga100% (1)
- Biologia Unad Trabajo FinalDokument8 SeitenBiologia Unad Trabajo FinalAMAIA CELESTE CHAMORRO FAJARDONoch keine Bewertungen
- SDFSDFSDDokument3 SeitenSDFSDFSDFabian CoronadoNoch keine Bewertungen
- Apoptosis Fisiología Animal 2023 Nutrición FisiologíaDokument41 SeitenApoptosis Fisiología Animal 2023 Nutrición FisiologíaCleoptrisLatemarginatusNoch keine Bewertungen
- Microbiologia ResumenDokument6 SeitenMicrobiologia ResumenSharon Daniela Ramirez CortesNoch keine Bewertungen
- Biologia 1Dokument100 SeitenBiologia 1Jhoana PeñaNoch keine Bewertungen
- Cuestionario Biologia 2Dokument12 SeitenCuestionario Biologia 2Hernandez Chavez DavidNoch keine Bewertungen
- Práctica 1 - Embriología y GenéticaDokument21 SeitenPráctica 1 - Embriología y GenéticaJHAJAYRA SHANTAL SEGURA MAZANoch keine Bewertungen
- Guion de EntrevistasDokument16 SeitenGuion de Entrevistasomaira gutierrezNoch keine Bewertungen
- Proteolisis Equipo 1Dokument10 SeitenProteolisis Equipo 1RAFAEL GONZÁLEZ VALENCIANoch keine Bewertungen
- Citoplasma Celular ResumenDokument3 SeitenCitoplasma Celular ResumenJessica Paulina Salazar RuizNoch keine Bewertungen
- Reporte de Práctica 8 Biología CelularDokument5 SeitenReporte de Práctica 8 Biología Celularvaledamian.004Noch keine Bewertungen
- Mecanica - Ciencia y Tecnologia U1 A01Dokument87 SeitenMecanica - Ciencia y Tecnologia U1 A01Alejandro CardozoNoch keine Bewertungen
- Calidad - Buenas Practicas OCDE (1) Aurora ZuzuarreguiDokument37 SeitenCalidad - Buenas Practicas OCDE (1) Aurora Zuzuarreguijulieth cNoch keine Bewertungen
- Cinética e Inhibición EnzimáticaDokument5 SeitenCinética e Inhibición EnzimáticaArévaloNoch keine Bewertungen
- Ejercicios 10 y 11 Del Tema 3Dokument1 SeiteEjercicios 10 y 11 Del Tema 3NEREA LAGUNA SERRANONoch keine Bewertungen
- Hibridación in Situ Con Fluorescencia PDFDokument7 SeitenHibridación in Situ Con Fluorescencia PDFValeria FigueroaNoch keine Bewertungen
- Historia de La Evolución de La CélulaDokument28 SeitenHistoria de La Evolución de La CélulaElena SalazarNoch keine Bewertungen
- Nanoarquitectura Medido Por andDokument10 SeitenNanoarquitectura Medido Por andlizbet MaiteNoch keine Bewertungen
- ENZIMASDokument5 SeitenENZIMASRoger Acosta MataNoch keine Bewertungen
- La Generación EspontáneaDokument2 SeitenLa Generación EspontáneaSarita Quispe AldereteNoch keine Bewertungen
- Tema 16Dokument8 SeitenTema 16Marta AlferezNoch keine Bewertungen
- Practica #03 de QuimicaDokument6 SeitenPractica #03 de QuimicaAnonymous LmL6HH1CRNoch keine Bewertungen
- Manualgeo Cap - 17 PDFDokument46 SeitenManualgeo Cap - 17 PDFEngineer6666Noch keine Bewertungen