Das könnte Ihnen auch gefallen
- Informe Practica 4 Grupo 1Dokument14 SeitenInforme Practica 4 Grupo 1Carlenis RamosNoch keine Bewertungen
- Practica de Bioquimica 4Dokument7 SeitenPractica de Bioquimica 4camilo100% (1)
- Equilibrio Gibbs DonnanDokument2 SeitenEquilibrio Gibbs DonnanDanielNoch keine Bewertungen
- Masa Proteica Visceral y EsqueléticaDokument7 SeitenMasa Proteica Visceral y EsqueléticaNicole RamirezNoch keine Bewertungen
- Discusion de Grupo N 3Dokument14 SeitenDiscusion de Grupo N 3RubenPerez100% (4)
- Metabolismo de Acidos Grasos.Dokument7 SeitenMetabolismo de Acidos Grasos.Mariel AybarNoch keine Bewertungen
- Practica N° 5 Fisiologia - OdtDokument6 SeitenPractica N° 5 Fisiologia - OdtVictor Lechuga Noa100% (1)
- Aislamiento de Linfocitos Mediante Centrifugación en Gradiente de DensidadDokument7 SeitenAislamiento de Linfocitos Mediante Centrifugación en Gradiente de DensidadJuan Pedro Rivas TorresNoch keine Bewertungen
- Manual NancyDokument10 SeitenManual NancynoemiNoch keine Bewertungen
- LCRDokument50 SeitenLCRalejandry100% (4)
- SINTESIS DE COLESTEROL ResumenDokument4 SeitenSINTESIS DE COLESTEROL ResumenXena Katherine Gutarra SosaNoch keine Bewertungen
- Clase 03 Agua Endogena y ExogenaDokument26 SeitenClase 03 Agua Endogena y ExogenaVictor Dimas Ramos Tapia0% (1)
- PRACTICA 06 - Metabolismo en Ratas 2021-2 VIRTUALDokument8 SeitenPRACTICA 06 - Metabolismo en Ratas 2021-2 VIRTUALAida SanchezNoch keine Bewertungen
- Difusion y Medio Ambiente InternoDokument25 SeitenDifusion y Medio Ambiente InternoAbner Chavez100% (2)
- Guía Práctica - Leyes de NewtonDokument4 SeitenGuía Práctica - Leyes de NewtonALEXIS FLORENCIO PICCININI MACEDONoch keine Bewertungen
- Practica 15Dokument8 SeitenPractica 15ALdo Aaron OR0% (1)
- Guia Laboratorio Glucogenólisis - PRÁCTICA No 8Dokument9 SeitenGuia Laboratorio Glucogenólisis - PRÁCTICA No 8Jessica Alexandra Peña MorenoNoch keine Bewertungen
- Practica N 06Dokument8 SeitenPractica N 06Maicol RojasNoch keine Bewertungen
- GLUCOGENOGENESISDokument2 SeitenGLUCOGENOGENESISMARGARITA MOO CHANNoch keine Bewertungen
- Folder de Trabajo - Semana 4 y 5Dokument16 SeitenFolder de Trabajo - Semana 4 y 5Pepe Rojas YachasNoch keine Bewertungen
- Practica Nº3 Cinetica EnzimaticaDokument30 SeitenPractica Nº3 Cinetica EnzimaticaClexis GonzalezNoch keine Bewertungen
- 5.2 Tabla de Vías MetabólicasDokument3 Seiten5.2 Tabla de Vías MetabólicasRebeca RioNoch keine Bewertungen
- Parciales Cat 1 r1 2do Cuatri Catedra 1Dokument29 SeitenParciales Cat 1 r1 2do Cuatri Catedra 1aleffrichNoch keine Bewertungen
- EXPERRIMENTO 13 Endogrino GrupalDokument16 SeitenEXPERRIMENTO 13 Endogrino GrupalSHEILA CESPEDESNoch keine Bewertungen
- Practica 8Dokument6 SeitenPractica 8Ysmael Barrientos100% (1)
- Electrolitos AURDokument40 SeitenElectrolitos AURAlexander VillaNoch keine Bewertungen
- Rutas Metabólicas - Leones - Por - La - SaludDokument25 SeitenRutas Metabólicas - Leones - Por - La - SaludALAN FERNANDO GONZALEZ SERRANONoch keine Bewertungen
- Unidad IV. Alcoholes Fenoles Eteres y Compuestos RelacionadosDokument49 SeitenUnidad IV. Alcoholes Fenoles Eteres y Compuestos RelacionadosFabiana DazaNoch keine Bewertungen
- PRACTICA 11creatininaDokument2 SeitenPRACTICA 11creatininaGERARDO SILVESTRE MELENDEZ MACALOPUNoch keine Bewertungen
- Final Bioquimica FMED UBA 15-12-17 TEMA ADokument2 SeitenFinal Bioquimica FMED UBA 15-12-17 TEMA ACande AmiconeNoch keine Bewertungen
- Metabolismo de La FructosaDokument9 SeitenMetabolismo de La FructosaYenniffer Vicente Maya0% (1)
- FOROSDokument18 SeitenFOROSNoelia VillaNoch keine Bewertungen
- Actividad 4 Ejercicio 3Dokument2 SeitenActividad 4 Ejercicio 3Harold Andia100% (1)
- Adme Del EtanolDokument18 SeitenAdme Del EtanolVanny RuizNoch keine Bewertungen
- CASO CLÍNICO No. 2Dokument15 SeitenCASO CLÍNICO No. 2Kenny MedranoNoch keine Bewertungen
- Metabolismo Del Glucógeno PDFDokument45 SeitenMetabolismo Del Glucógeno PDFJuly CamposNoch keine Bewertungen
- Etanol 09.Ppt 3ºDokument17 SeitenEtanol 09.Ppt 3ºRenzo FormiaNoch keine Bewertungen
- Enfermedades MonogénicasDokument31 SeitenEnfermedades MonogénicasDiabloNoch keine Bewertungen
- Mètodos para La Determinación de Proteínas Totales y Albúmina en SueroDokument16 SeitenMètodos para La Determinación de Proteínas Totales y Albúmina en SueroGino Chávez SánchezNoch keine Bewertungen
- Determinacion de Glucogeno en Higado y CorazonDokument2 SeitenDeterminacion de Glucogeno en Higado y Corazonhernando guerreroNoch keine Bewertungen
- Acumulaciones IntracelularesDokument2 SeitenAcumulaciones IntracelularesRolfyrasdvgjpuweg io p GuzmanNoch keine Bewertungen
- Deficiencia de La Glucosa-6-Fosfato Deshidrogenasa (G6PD)Dokument13 SeitenDeficiencia de La Glucosa-6-Fosfato Deshidrogenasa (G6PD)medicinaensNoch keine Bewertungen
- Esofago y EstomagoDokument6 SeitenEsofago y EstomagoCarlos ValenzuelaNoch keine Bewertungen
- Lab. Bioqui Semana 10Dokument5 SeitenLab. Bioqui Semana 10Aylin ArellanoNoch keine Bewertungen
- Cuestionario - Tema 10Dokument5 SeitenCuestionario - Tema 10María del Carmen Ramírez SalinasNoch keine Bewertungen
- PRÁCTICA #08 EnzimasDokument6 SeitenPRÁCTICA #08 Enzimasjimmy gomezNoch keine Bewertungen
- Guia de Practicas 2017-I UnwDokument78 SeitenGuia de Practicas 2017-I UnwAnonymous q3hUTA0% (1)
- Practica 1Dokument9 SeitenPractica 1janssenNoch keine Bewertungen
- Práctica #1 de Laboratorio Fisiología Neuromuscular y de Los Sistemas Sensoriales 2016Dokument6 SeitenPráctica #1 de Laboratorio Fisiología Neuromuscular y de Los Sistemas Sensoriales 2016Carlos Aquise LaricoNoch keine Bewertungen
- Guia 2023-2-FinalDokument67 SeitenGuia 2023-2-FinalMilagros A0% (1)
- Informe de Práctica #9 - LABORATORIO BIOQUIMICA I - 2022Dokument9 SeitenInforme de Práctica #9 - LABORATORIO BIOQUIMICA I - 2022Ursula Asty100% (1)
- Póster Glucólisis AnaeróbicaDokument1 SeitePóster Glucólisis AnaeróbicaNaomi SacaNoch keine Bewertungen
- 4 InformeDokument11 Seiten4 InformeJoseth De Jesus Larios CortezNoch keine Bewertungen
- Informe 5Dokument20 SeitenInforme 5Isabella IturreyNoch keine Bewertungen
- Bioquimica Informe Ayuna NosotrosDokument12 SeitenBioquimica Informe Ayuna NosotrosGeorman Dantas Leon100% (1)
- INTEROGANTES BioquimicaDokument11 SeitenINTEROGANTES Bioquimicafabripro2Noch keine Bewertungen
- Práctica N°8 Glucógeno Hepático Grupo'09 (1) BBHBDokument9 SeitenPráctica N°8 Glucógeno Hepático Grupo'09 (1) BBHBluianvil3Noch keine Bewertungen
- Practica N 6Dokument7 SeitenPractica N 6Irma NeyraNoch keine Bewertungen
- Practica 5 Metabolismo Intermediario. EfDokument15 SeitenPractica 5 Metabolismo Intermediario. EfMariajose BolivarNoch keine Bewertungen
- Practica N 4Dokument4 SeitenPractica N 4Yuliet Madyori BautistaNoch keine Bewertungen
- Horario 5to 2019 PDFDokument1 SeiteHorario 5to 2019 PDFUzziel LyonNoch keine Bewertungen
- Constinscripcion21175380 PDFDokument1 SeiteConstinscripcion21175380 PDFSergio Hernandez El ReyNoch keine Bewertungen
- Horario 5to 2019 PDFDokument1 SeiteHorario 5to 2019 PDFUzziel LyonNoch keine Bewertungen
- Has Dejado Tu Primer AmorDokument7 SeitenHas Dejado Tu Primer AmorUzziel Lyon100% (1)
- Resurreccion de CristoDokument3 SeitenResurreccion de CristoUzziel LyonNoch keine Bewertungen
- 1 Cor 11 (17-34) Unidad Del CuerpoDokument3 Seiten1 Cor 11 (17-34) Unidad Del CuerpoUzziel LyonNoch keine Bewertungen
- Los 5 Lenguajes Del AmorDokument11 SeitenLos 5 Lenguajes Del AmorUzziel LyonNoch keine Bewertungen
- Equilibrio RedoxDokument11 SeitenEquilibrio RedoxIvan JimenezNoch keine Bewertungen
- Tramites A Realizar Antes de Viajar Al Exterio PastorDokument5 SeitenTramites A Realizar Antes de Viajar Al Exterio PastorUzziel LyonNoch keine Bewertungen
- Tabla de Los TemperamentosDokument2 SeitenTabla de Los TemperamentosUzziel LyonNoch keine Bewertungen
- Visa Responsabilidad Democrática Chile PDFDokument3 SeitenVisa Responsabilidad Democrática Chile PDFUzziel LyonNoch keine Bewertungen
- Sayjan Lus de - Magia Sexual Positiva PDFDokument67 SeitenSayjan Lus de - Magia Sexual Positiva PDFAndita100% (4)
- Desarrollo Actividad 2 PasteleriaDokument4 SeitenDesarrollo Actividad 2 PasteleriaCarlos Adonnai CardenasNoch keine Bewertungen
- Práctica 11 - Dilatación de SólidosDokument3 SeitenPráctica 11 - Dilatación de SólidosSAUL BOLIVAR SARMIENTO CARCHINoch keine Bewertungen
- Pelvis PpsDokument21 SeitenPelvis PpsJj GoNoch keine Bewertungen
- Hernia Inguinal 2.Dokument10 SeitenHernia Inguinal 2.Nathaly MontoyaNoch keine Bewertungen
- Evaluacion Naturales 5 U5Dokument3 SeitenEvaluacion Naturales 5 U5Bely de CasasNoch keine Bewertungen
- Examen La Celula BiologiaDokument3 SeitenExamen La Celula Biologiamaira alejandra muñoz0% (1)
- MuestreoDokument23 SeitenMuestreoAnyi Paola Martinez RicardoNoch keine Bewertungen
- Informe - de - Practicas Ubap Metropolitano ChimboteDokument43 SeitenInforme - de - Practicas Ubap Metropolitano ChimboteCarlos Jose Peje QuesadaNoch keine Bewertungen
- AntibióticosDokument28 SeitenAntibióticoslalore07Noch keine Bewertungen
- Revision de Andamios Formato ModeloDokument2 SeitenRevision de Andamios Formato ModelocesarleonelcortezNoch keine Bewertungen
- Blanqueo AlgodonDokument16 SeitenBlanqueo AlgodonGonzalo Jara PacoriNoch keine Bewertungen
- Material de Estudio - KINESIOLOGIA PACIENTES ONCOLOGICOSDokument60 SeitenMaterial de Estudio - KINESIOLOGIA PACIENTES ONCOLOGICOSNicol Sandoval Soto100% (1)
- El Concepto de SuperyoDokument21 SeitenEl Concepto de SuperyoRosa María López100% (1)
- Ideologias y Nuevos ConflictosDokument7 SeitenIdeologias y Nuevos ConflictosSamuel SolisNoch keine Bewertungen
- Bases Físicas de La EcografíaDokument79 SeitenBases Físicas de La Ecografíaterebgarcia100% (2)
- Mapa Conceptual de La AutoestimaDokument1 SeiteMapa Conceptual de La AutoestimaNorma RamiresNoch keine Bewertungen
- 1 - LA PUBERTAD Y LA ADOLESCENCIA (Delval, Juan)Dokument11 Seiten1 - LA PUBERTAD Y LA ADOLESCENCIA (Delval, Juan)Eliud Guerrero100% (1)
- Guía N°1 - Introducción A La Transferencia de Calor PDFDokument2 SeitenGuía N°1 - Introducción A La Transferencia de Calor PDFDanny GomezNoch keine Bewertungen
- TESIS Tipos y Características de Tuberías para Elaboración de Pozos PetrolerosDokument190 SeitenTESIS Tipos y Características de Tuberías para Elaboración de Pozos PetrolerosAlexis Michelle Roca100% (3)
- Guion LiterarioDokument2 SeitenGuion LiterarioAracelis PaoliniNoch keine Bewertungen
- FILA A Prueba Nomenclatura InorganicaDokument5 SeitenFILA A Prueba Nomenclatura InorganicaANA MARIA MARIHUAN100% (1)
- Reproducción de MMS y MMLDokument3 SeitenReproducción de MMS y MMLEnrique Llano BernalNoch keine Bewertungen
- Curso CPR Jerez MAsajes3Dokument68 SeitenCurso CPR Jerez MAsajes3mariabethlemNoch keine Bewertungen
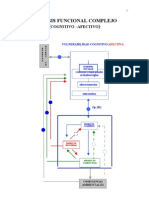
- Walter Riso - Análisis Funcional ComplejoDokument8 SeitenWalter Riso - Análisis Funcional ComplejoSamantha Rojas GuzmanNoch keine Bewertungen
- ContaminacionDokument3 SeitenContaminacionJosé Armando Orizano AlaniaNoch keine Bewertungen
- Ingenuity 128Dokument7 SeitenIngenuity 128Anonymous F1xtU2RNoch keine Bewertungen
- Fresas PDFDokument114 SeitenFresas PDFJose MarquezNoch keine Bewertungen
- Akron Hydraulic XLF 32, XLF 46, XLF 68Dokument7 SeitenAkron Hydraulic XLF 32, XLF 46, XLF 68aramirezharoNoch keine Bewertungen
- Universidad Regional Autónoma de Los Andes UniandesDokument23 SeitenUniversidad Regional Autónoma de Los Andes UniandesCarballo GiovannyNoch keine Bewertungen