Das könnte Ihnen auch gefallen
- Ma2 PDFDokument3 SeitenMa2 PDFJoyceline DavidNoch keine Bewertungen
- 2013 SLAF-seq - An Efficient Method of Large-Scale de Novo SNP Discovery and Genotyping Using High-Throughput SequencingDokument9 Seiten2013 SLAF-seq - An Efficient Method of Large-Scale de Novo SNP Discovery and Genotyping Using High-Throughput Sequencingwsx kNoch keine Bewertungen
- Application of Biotechnology To Wheat ImprovementDokument28 SeitenApplication of Biotechnology To Wheat ImprovementJagadeeshNoch keine Bewertungen
- Mol Breed - 1 - 7 - MergedDokument88 SeitenMol Breed - 1 - 7 - MergedSukanta RoyNoch keine Bewertungen
- Mol Breed - 1Dokument21 SeitenMol Breed - 1Sukanta RoyNoch keine Bewertungen
- Molecular Markers: Their Use in Tree Improvement: R. Mahajan, P. GuptaDokument8 SeitenMolecular Markers: Their Use in Tree Improvement: R. Mahajan, P. GuptaAbdul basitNoch keine Bewertungen
- 1 s2.0 S2405844021021964 MainDokument20 Seiten1 s2.0 S2405844021021964 MainKERTÉSZMÉRNÖK SZIEMKKNoch keine Bewertungen
- A Method For Discovery of Genome-Wide SNP Between Any Two Genotypes From Whole-Genome Re-Sequencing DataDokument8 SeitenA Method For Discovery of Genome-Wide SNP Between Any Two Genotypes From Whole-Genome Re-Sequencing DataSekste MaludaNoch keine Bewertungen
- Brugmans Et Al. 2002 (Transcript Mapping)Dokument12 SeitenBrugmans Et Al. 2002 (Transcript Mapping)Sathish KumarNoch keine Bewertungen
- DNA Based Techniques For Studying Genetic DiversityDokument30 SeitenDNA Based Techniques For Studying Genetic DiversitySyahrul Tuba Al FatihNoch keine Bewertungen
- The Power of Genomics in AgricultureDokument5 SeitenThe Power of Genomics in Agriculturesumon khanNoch keine Bewertungen
- 12 OkayDokument13 Seiten12 Okayel8zaaNoch keine Bewertungen
- Genetic DiagnosaDokument6 SeitenGenetic DiagnosaIfany oschaliaNoch keine Bewertungen
- Applications of Molecular Markers and Genomic Tools in Pearl Millet (Cenchrus Americanus (L.) Morronesyn. Pennisetum Glaucum (L.) R. BR.)Dokument10 SeitenApplications of Molecular Markers and Genomic Tools in Pearl Millet (Cenchrus Americanus (L.) Morronesyn. Pennisetum Glaucum (L.) R. BR.)IJAR JOURNALNoch keine Bewertungen
- A Review On SNP and Other Types of Molecular Markers and Their Use in Animal GeneticsDokument31 SeitenA Review On SNP and Other Types of Molecular Markers and Their Use in Animal GeneticsboboyoyotNoch keine Bewertungen
- Aflp Genotyping and Fingerprinting: Ulrich G. Mueller and L. Lareesa WolfenbargerDokument6 SeitenAflp Genotyping and Fingerprinting: Ulrich G. Mueller and L. Lareesa WolfenbargerHelpBiotech IndiaNoch keine Bewertungen
- DNA Finger PrintingDokument21 SeitenDNA Finger PrintingMeylinda EnggiNoch keine Bewertungen
- Applications of Bulking in Molecular Characterization of Plant Germplasm A Critical ReviewDokument7 SeitenApplications of Bulking in Molecular Characterization of Plant Germplasm A Critical ReviewIrena CalicNoch keine Bewertungen
- Accessing Genetic Variation: Genotyping Single Nucleotide PolymorphismsDokument13 SeitenAccessing Genetic Variation: Genotyping Single Nucleotide PolymorphismsVicky G OeiNoch keine Bewertungen
- Kumar Jai Anand Doctral SeminarDokument32 SeitenKumar Jai Anand Doctral Seminarkumar jaianandNoch keine Bewertungen
- Molecular Markers in Plant Genome AnalysisDokument13 SeitenMolecular Markers in Plant Genome Analysisarunnene2Noch keine Bewertungen
- Whole-Genome Resequencing: Changing The Paradigms of SNP Detection, Molecular Mapping and Gene DiscoveryDokument11 SeitenWhole-Genome Resequencing: Changing The Paradigms of SNP Detection, Molecular Mapping and Gene DiscoverySekste MaludaNoch keine Bewertungen
- DNA Based Molecular MarkersDokument34 SeitenDNA Based Molecular MarkersBlaxez YTNoch keine Bewertungen
- Bulletin 6004 HRM GenotypingDokument6 SeitenBulletin 6004 HRM GenotypingFelipe Andres Contreras SepulvedaNoch keine Bewertungen
- Marcadores SNP y El Impacto en El FitomejoramientoDokument12 SeitenMarcadores SNP y El Impacto en El FitomejoramientoDayani GomezNoch keine Bewertungen
- Bioinformatics TM3Dokument26 SeitenBioinformatics TM3subhanilchakrabortyNoch keine Bewertungen
- Agarwal 2008 - Advances in Molecular Marker Techniques and Their Applications in Plant SciencesDokument16 SeitenAgarwal 2008 - Advances in Molecular Marker Techniques and Their Applications in Plant SciencessherenelaiNoch keine Bewertungen
- Prediction-of-Sorghum-bicolor-Genotype-from In-situ-Images-UsingDokument9 SeitenPrediction-of-Sorghum-bicolor-Genotype-from In-situ-Images-Usingtagne simo rodrigueNoch keine Bewertungen
- c2Dokument37 Seitenc2Alka GuptaNoch keine Bewertungen
- Simple Sequence RepeatsDokument1 SeiteSimple Sequence Repeatsmilove_88003272Noch keine Bewertungen
- Molecular MarkersDokument42 SeitenMolecular Markerssaroj Burlakoti100% (1)
- R. R. Dhutmal, Et AlDokument10 SeitenR. R. Dhutmal, Et AlFernando Castro EchavezNoch keine Bewertungen
- Types of MarkersDokument6 SeitenTypes of MarkersAdinath MahipalNoch keine Bewertungen
- Comparison of SNP and CAPS Markers Application in Genetic Research in Wheat and BarleyDokument5 SeitenComparison of SNP and CAPS Markers Application in Genetic Research in Wheat and BarleyAlangelico Ortiz San PascualNoch keine Bewertungen
- Varshney Et Al. 2005 Genic MicrossatelliteDokument9 SeitenVarshney Et Al. 2005 Genic MicrossatelliteGisele HolandaNoch keine Bewertungen
- Chapter 12Dokument16 SeitenChapter 12Zeeshan UsmaniNoch keine Bewertungen
- Teknik/metode Analisis Keragaman Genetik/populasi/spesies: Dosen Pengampu Dr. Asmi Citra Malina, S.Pi, M.AgrDokument18 SeitenTeknik/metode Analisis Keragaman Genetik/populasi/spesies: Dosen Pengampu Dr. Asmi Citra Malina, S.Pi, M.AgrMuhammad Arif FauzanNoch keine Bewertungen
- Genetic Polymorphism and EvolutionDokument18 SeitenGenetic Polymorphism and EvolutionchatoyantNoch keine Bewertungen
- N.K Paper-1 Published PDFDokument7 SeitenN.K Paper-1 Published PDFNawal Kishor YadavNoch keine Bewertungen
- PDA-Global Oct 2011 Poster MR2Dokument1 SeitePDA-Global Oct 2011 Poster MR2sblazej7118Noch keine Bewertungen
- Plant Breeding - 2021 - Zhang - Development of Codominant SCAR Markers To Detect The Pto Tm22 I3 and Sw5 Genes in Tomato PDFDokument7 SeitenPlant Breeding - 2021 - Zhang - Development of Codominant SCAR Markers To Detect The Pto Tm22 I3 and Sw5 Genes in Tomato PDFKERTÉSZMÉRNÖK SZIEMKKNoch keine Bewertungen
- Quantitative Trait Loci (QTL) Mapping: Gurbachan S. MiglaniDokument42 SeitenQuantitative Trait Loci (QTL) Mapping: Gurbachan S. Miglanianon_310555357Noch keine Bewertungen
- Introduction To Molecular MarkersDokument47 SeitenIntroduction To Molecular MarkersSama MouradNoch keine Bewertungen
- NG 3624Dokument9 SeitenNG 3624Marcus TutertNoch keine Bewertungen
- Proposal Presentation Sea CucumberDokument11 SeitenProposal Presentation Sea CucumberNasrul Safwan MTNoch keine Bewertungen
- MAS in Sorghum Using KASP - 2019Dokument10 SeitenMAS in Sorghum Using KASP - 2019raveendra.gmNoch keine Bewertungen
- Molecular Markers in Forest Management and Tree Breeding: A ReviewDokument7 SeitenMolecular Markers in Forest Management and Tree Breeding: A ReviewFernando Castro EchavezNoch keine Bewertungen
- Journal of Molecular Biomarkers & Diagnosis: Molecular Markers: An Excellent Tool For Genetic AnalysisDokument2 SeitenJournal of Molecular Biomarkers & Diagnosis: Molecular Markers: An Excellent Tool For Genetic AnalysisfryddaproblesNoch keine Bewertungen
- SNaPshot® MultiplexDokument4 SeitenSNaPshot® MultiplexCrist VillarNoch keine Bewertungen
- Chapter 3Dokument15 SeitenChapter 3Stephani CortesNoch keine Bewertungen
- SNP 161121140004 PDFDokument24 SeitenSNP 161121140004 PDFDr-Dalya ShakirNoch keine Bewertungen
- SNP 161121140004 PDFDokument24 SeitenSNP 161121140004 PDFDr-Dalya ShakirNoch keine Bewertungen
- Transcriptome Sequencing of A Large Human Family Identifies The Impact of Rare Noncoding VariantsDokument12 SeitenTranscriptome Sequencing of A Large Human Family Identifies The Impact of Rare Noncoding VariantsJacintoOrozcoSamuelNoch keine Bewertungen
- 1471 2148-7-135 S2 SelectionofcandidatecodingDNADokument11 Seiten1471 2148-7-135 S2 SelectionofcandidatecodingDNAOanaNoch keine Bewertungen
- Molecular Quantitative Genetics For Plant Breeding Roundtable 2010xDokument69 SeitenMolecular Quantitative Genetics For Plant Breeding Roundtable 2010xMona ElsayedNoch keine Bewertungen
- 21 - Kebere Bezaweletaw - FinalPaperDokument11 Seiten21 - Kebere Bezaweletaw - FinalPaperiisteNoch keine Bewertungen
- 03 Marker Aided SelectionDokument45 Seiten03 Marker Aided Selectionanon_310555357100% (1)
- Clinical Molecular Diagnostic Techniques: A Brief ReviewDokument19 SeitenClinical Molecular Diagnostic Techniques: A Brief ReviewSorin LazarNoch keine Bewertungen
- Cytological Markers in Genome MappingDokument8 SeitenCytological Markers in Genome MappingInternational Journal of Innovative Science and Research Technology100% (1)
- Introduction to Bioinformatics, Sequence and Genome AnalysisVon EverandIntroduction to Bioinformatics, Sequence and Genome AnalysisNoch keine Bewertungen
- B. Geoinformatics PDFDokument77 SeitenB. Geoinformatics PDFmchakra720% (1)
- Opc PPT FinalDokument22 SeitenOpc PPT FinalnischalaNoch keine Bewertungen
- C C C C: "P P P P PDokument25 SeitenC C C C: "P P P P PShalu Dua KatyalNoch keine Bewertungen
- Companyprofil E: Erfanconstructionsolut IonDokument14 SeitenCompanyprofil E: Erfanconstructionsolut IonNurin AleesyaNoch keine Bewertungen
- MASONRYDokument8 SeitenMASONRYJowelyn MaderalNoch keine Bewertungen
- Functional DesignDokument17 SeitenFunctional DesignRajivSharmaNoch keine Bewertungen
- Köppen Climate Classification - Wikipedia, The Free EncyclopediaDokument15 SeitenKöppen Climate Classification - Wikipedia, The Free EncyclopediaAndreea Tataru StanciNoch keine Bewertungen
- Iec TR 61010-3-020-1999Dokument76 SeitenIec TR 61010-3-020-1999Vasko MandilNoch keine Bewertungen
- Chronic Kidney DiseaseDokument15 SeitenChronic Kidney Diseaseapi-270623039Noch keine Bewertungen
- Mark Garside Resume May 2014Dokument3 SeitenMark Garside Resume May 2014api-199955558Noch keine Bewertungen
- 04 - Fetch Decode Execute Cycle PDFDokument3 Seiten04 - Fetch Decode Execute Cycle PDFShaun HaxaelNoch keine Bewertungen
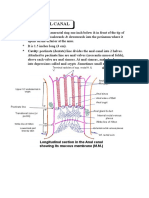
- Anatomy Anal CanalDokument14 SeitenAnatomy Anal CanalBela Ronaldoe100% (1)
- Costbenefit Analysis 2015Dokument459 SeitenCostbenefit Analysis 2015TRÂM NGUYỄN THỊ BÍCHNoch keine Bewertungen
- LM74680 Fasson® Fastrans NG Synthetic PE (ST) / S-2050/ CK40Dokument2 SeitenLM74680 Fasson® Fastrans NG Synthetic PE (ST) / S-2050/ CK40Nishant JhaNoch keine Bewertungen
- Aptitude Number System PDFDokument5 SeitenAptitude Number System PDFharieswaranNoch keine Bewertungen
- Duavent Drug Study - CunadoDokument3 SeitenDuavent Drug Study - CunadoLexa Moreene Cu�adoNoch keine Bewertungen
- Acute Coronary SyndromeDokument30 SeitenAcute Coronary SyndromeEndar EszterNoch keine Bewertungen
- A Case Study of Coustomer Satisfaction in Demat Account At: A Summer Training ReportDokument110 SeitenA Case Study of Coustomer Satisfaction in Demat Account At: A Summer Training ReportDeepak SinghalNoch keine Bewertungen
- Playful Homeschool Planner - FULLDokument13 SeitenPlayful Homeschool Planner - FULLamandalecuyer88Noch keine Bewertungen
- Outdoor Air Pollution: Sources, Health Effects and SolutionsDokument20 SeitenOutdoor Air Pollution: Sources, Health Effects and SolutionsCamelia RadulescuNoch keine Bewertungen
- Sindi and Wahab in 18th CenturyDokument9 SeitenSindi and Wahab in 18th CenturyMujahid Asaadullah AbdullahNoch keine Bewertungen
- Presentation 11Dokument14 SeitenPresentation 11stellabrown535Noch keine Bewertungen
- of Thesis ProjectDokument2 Seitenof Thesis ProjectmoonNoch keine Bewertungen
- The Magic DrumDokument185 SeitenThe Magic Drumtanishgiri2012Noch keine Bewertungen
- Project ManagementDokument11 SeitenProject ManagementBonaventure NzeyimanaNoch keine Bewertungen
- Unit 7: Anthropology: Q2e Listening & Speaking 4: Audio ScriptDokument6 SeitenUnit 7: Anthropology: Q2e Listening & Speaking 4: Audio ScriptĐại học Bạc Liêu Truyền thông100% (1)
- Mixed Up MonstersDokument33 SeitenMixed Up MonstersjaneNoch keine Bewertungen
- Bana LingaDokument9 SeitenBana LingaNimai Pandita Raja DasaNoch keine Bewertungen
- What Is TranslationDokument3 SeitenWhat Is TranslationSanskriti MehtaNoch keine Bewertungen
- My Personal Code of Ethics1Dokument1 SeiteMy Personal Code of Ethics1Princess Angel LucanasNoch keine Bewertungen