Das könnte Ihnen auch gefallen
- AMINOÁCIDOSDokument11 SeitenAMINOÁCIDOSHeriberto AngelesNoch keine Bewertungen
- ColesterolDokument54 SeitenColesterolStalin PintoNoch keine Bewertungen
- Ejercicios de Tablas de Frecuencias y Gráficos-AliciaDokument16 SeitenEjercicios de Tablas de Frecuencias y Gráficos-AliciaAlicia Chavez CernaNoch keine Bewertungen
- Diferenciación CelularDokument7 SeitenDiferenciación CelularAlicia Chavez CernaNoch keine Bewertungen
- Técnicas en Bioquímica y Biología MolecularDokument2 SeitenTécnicas en Bioquímica y Biología MolecularAlicia Chavez CernaNoch keine Bewertungen
- Anatomía Del Túnel Del CarpoDokument2 SeitenAnatomía Del Túnel Del CarpoAlicia Chavez CernaNoch keine Bewertungen
- Diferenciación CelularDokument6 SeitenDiferenciación CelularAlicia Chavez CernaNoch keine Bewertungen
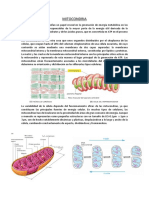
- La MitocondriaDokument11 SeitenLa MitocondriaAlicia Chavez CernaNoch keine Bewertungen
- Informe Medidas y ProliferacionDokument5 SeitenInforme Medidas y ProliferacionAlicia Chavez CernaNoch keine Bewertungen
- Vasos LinfáticosDokument5 SeitenVasos LinfáticosAlicia Chavez CernaNoch keine Bewertungen
- Tarea I - Ejercicios Del 1 Al 7Dokument8 SeitenTarea I - Ejercicios Del 1 Al 7Alicia Chavez CernaNoch keine Bewertungen
- Túnica Íntima, Media y Adventicia.. DiapositivasDokument5 SeitenTúnica Íntima, Media y Adventicia.. DiapositivasAlicia Chavez CernaNoch keine Bewertungen
- Región SuboptoestriadaDokument5 SeitenRegión SuboptoestriadaAlicia Chavez CernaNoch keine Bewertungen
- Tarea I - Ejercicios Del 1 Al 7Dokument8 SeitenTarea I - Ejercicios Del 1 Al 7Alicia Chavez CernaNoch keine Bewertungen
- Hemangiomas: Una RevisiónDokument4 SeitenHemangiomas: Una RevisiónAlicia Chavez CernaNoch keine Bewertungen
- List No 1Dokument4 SeitenList No 1Alicia Chavez CernaNoch keine Bewertungen
- La EugenesiaDokument5 SeitenLa EugenesiaAlicia Chavez CernaNoch keine Bewertungen
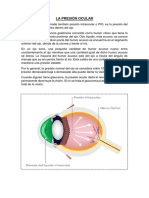
- El Humor AcuosoDokument5 SeitenEl Humor AcuosoAlicia Chavez CernaNoch keine Bewertungen
- Síndrome Del Túnel CarpianoDokument4 SeitenSíndrome Del Túnel CarpianoAlicia Chavez CernaNoch keine Bewertungen
- La Colonia en La Historia Del Derecho PeruanoDokument1 SeiteLa Colonia en La Historia Del Derecho PeruanoAlicia Chavez CernaNoch keine Bewertungen
- 05-16 2 PDFDokument10 Seiten05-16 2 PDFMaria L. FuentesNoch keine Bewertungen
- Facial AnastomosisDokument3 SeitenFacial AnastomosisAlicia Chavez CernaNoch keine Bewertungen
- Sistemas de Grupos SanguineosDokument25 SeitenSistemas de Grupos SanguineosMarcelo BecksNoch keine Bewertungen
- Respiracion CelularDokument24 SeitenRespiracion CelularRicardo HernándezNoch keine Bewertungen
- Tarea 1 - AnticuerposDokument2 SeitenTarea 1 - Anticuerposcristian pulidoNoch keine Bewertungen
- Composición Química de Los Seres VivosDokument1 SeiteComposición Química de Los Seres VivosPatrick Mantilla Bacilio100% (1)
- Trip To FanoDokument28 SeitenTrip To FanoAlexandra AlvarezNoch keine Bewertungen
- Folleto Suplementos Mondragon-1Dokument4 SeitenFolleto Suplementos Mondragon-1Tonatiuh VelázquezNoch keine Bewertungen
- Codigo Genetico EjerciciosDokument6 SeitenCodigo Genetico EjerciciosIrina Castrillo CorralesNoch keine Bewertungen
- Estructura y Función Del ADN. Información ComplementariaDokument13 SeitenEstructura y Función Del ADN. Información ComplementariaAnairis MendozaNoch keine Bewertungen
- Actividad 2. Cuantificación de ProteínasDokument4 SeitenActividad 2. Cuantificación de ProteínasYadra GascaNoch keine Bewertungen
- Ácidos NucleicosDokument15 SeitenÁcidos NucleicosMartín GamarraNoch keine Bewertungen
- BiomoleculasDokument7 SeitenBiomoleculasRonnie GenezNoch keine Bewertungen
- Complejo Multienzimático Que Interviene en La Biosíntesis de Ácidos GrasosDokument3 SeitenComplejo Multienzimático Que Interviene en La Biosíntesis de Ácidos GrasosMichaelRvNoch keine Bewertungen
- Informe de Quimica NantarDokument7 SeitenInforme de Quimica Nantarmaria belenNoch keine Bewertungen
- Propiedades de La SardinaDokument21 SeitenPropiedades de La SardinaInternet Magia AzulNoch keine Bewertungen
- Biología 3 UNMSMDokument12 SeitenBiología 3 UNMSMJunnior PINZAS AYALANoch keine Bewertungen
- Lípidos, Ácidos Grasos y TriacilglicerolesDokument19 SeitenLípidos, Ácidos Grasos y TriacilglicerolesDante SegoviaNoch keine Bewertungen
- Bioquímica 15 Enzimología y Bioenergética Nilo ChicaDokument2 SeitenBioquímica 15 Enzimología y Bioenergética Nilo Chicaludwing1234Noch keine Bewertungen
- Logica CuantificacionalDokument27 SeitenLogica CuantificacionalSheyla Castillo100% (1)
- 3 Replicación de ADN. Grupo2Dokument27 Seiten3 Replicación de ADN. Grupo2July Aguila100% (1)
- Blgo. Ms. Pablo Chuna MogollónDokument24 SeitenBlgo. Ms. Pablo Chuna MogollónAnonymous aeyMBcNDfNoch keine Bewertungen
- Cocteles Mas Usados MESOESTETIC PDFDokument3 SeitenCocteles Mas Usados MESOESTETIC PDFCarlos casayasNoch keine Bewertungen
- Ácidos Nucleicos (Modo de Compatibilidad)Dokument114 SeitenÁcidos Nucleicos (Modo de Compatibilidad)Alejo Diez PatiñoNoch keine Bewertungen
- Mapa Conceptual de Las Proteinas 4Dokument1 SeiteMapa Conceptual de Las Proteinas 4Maite DiscorNoch keine Bewertungen
- ChaperonasDokument62 SeitenChaperonasEsteban RobloxNoch keine Bewertungen
- Seminario 2Dokument6 SeitenSeminario 2Mireia PenaNoch keine Bewertungen
- Factores Que Afectan La Actividad EnzimáticaDokument6 SeitenFactores Que Afectan La Actividad EnzimáticaJosué MNoch keine Bewertungen
- Mapa Conceptual CarbohidratosDokument1 SeiteMapa Conceptual CarbohidratosCatherine Sofia Ochoa BarriosNoch keine Bewertungen
- Metabolismo de Acidos NucleicosDokument30 SeitenMetabolismo de Acidos NucleicosHernàn RV0% (1)