Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- 2009 2011 DS Manual - Club Car (001-061)Dokument61 Seiten2009 2011 DS Manual - Club Car (001-061)misaNoch keine Bewertungen
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Analysis of Electric Machinery Krause Manual Solution PDFDokument2 SeitenAnalysis of Electric Machinery Krause Manual Solution PDFKuldeep25% (8)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- CH 1 - Democracy and American PoliticsDokument9 SeitenCH 1 - Democracy and American PoliticsAndrew Philip ClarkNoch keine Bewertungen
- Dokumen - Pub - Bobs Refunding Ebook v3 PDFDokument65 SeitenDokumen - Pub - Bobs Refunding Ebook v3 PDFJohn the First100% (3)
- Lesson 5 Designing and Developing Social AdvocacyDokument27 SeitenLesson 5 Designing and Developing Social Advocacydaniel loberizNoch keine Bewertungen
- Project Scheduling and TrackingDokument47 SeitenProject Scheduling and TrackingArun VinodhNoch keine Bewertungen
- Modulo EminicDokument13 SeitenModulo EminicAndreaNoch keine Bewertungen
- Mark Garside Resume May 2014Dokument3 SeitenMark Garside Resume May 2014api-199955558Noch keine Bewertungen
- Mcom Sem 4 Project FinalDokument70 SeitenMcom Sem 4 Project Finallaxmi iyer75% (4)
- Week 7Dokument24 SeitenWeek 7Priyank PatelNoch keine Bewertungen
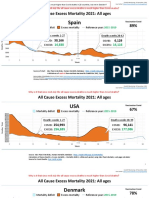
- Countries EXCESS DEATHS All Ages - 15nov2021Dokument21 SeitenCountries EXCESS DEATHS All Ages - 15nov2021robaksNoch keine Bewertungen
- An Exploration of The Ethno-Medicinal Practices Among Traditional Healers in Southwest Cebu, PhilippinesDokument7 SeitenAn Exploration of The Ethno-Medicinal Practices Among Traditional Healers in Southwest Cebu, PhilippinesleecubongNoch keine Bewertungen
- Ozone Therapy - A Clinical Review A. M. Elvis and J. S. EktaDokument5 SeitenOzone Therapy - A Clinical Review A. M. Elvis and J. S. Ektatahuti696Noch keine Bewertungen
- MGMT Audit Report WritingDokument28 SeitenMGMT Audit Report WritingAndrei IulianNoch keine Bewertungen
- Linguistics Is Descriptive, Not Prescriptive.: Prescriptive Grammar. Prescriptive Rules Tell You HowDokument2 SeitenLinguistics Is Descriptive, Not Prescriptive.: Prescriptive Grammar. Prescriptive Rules Tell You HowMonette Rivera Villanueva100% (1)
- Crown WF-3000 1.2Dokument5 SeitenCrown WF-3000 1.2Qirat KhanNoch keine Bewertungen
- DP 2 Human IngenuityDokument8 SeitenDP 2 Human Ingenuityamacodoudiouf02Noch keine Bewertungen
- Chapter3 Elasticity and ForecastingDokument25 SeitenChapter3 Elasticity and ForecastingGee JoeNoch keine Bewertungen
- Article An Incident and Injury Free Culture Changing The Face of Project Operations Terra117 2Dokument6 SeitenArticle An Incident and Injury Free Culture Changing The Face of Project Operations Terra117 2nguyenthanhtuan_ecoNoch keine Bewertungen
- Post Appraisal InterviewDokument3 SeitenPost Appraisal InterviewNidhi D100% (1)
- Floating Oil Skimmer Design Using Rotary Disc MethDokument9 SeitenFloating Oil Skimmer Design Using Rotary Disc MethAhmad YaniNoch keine Bewertungen
- De Thi Hoc Ki 1 Mon Tieng Anh Lop 5 Co File NgheDokument10 SeitenDe Thi Hoc Ki 1 Mon Tieng Anh Lop 5 Co File Nghetuyen truongNoch keine Bewertungen
- Turning PointsDokument2 SeitenTurning Pointsapi-223780825Noch keine Bewertungen
- CX Programmer Operation ManualDokument536 SeitenCX Programmer Operation ManualVefik KaraegeNoch keine Bewertungen
- Chemistry Form 4 Daily Lesson Plan - CompressDokument3 SeitenChemistry Form 4 Daily Lesson Plan - Compressadila ramlonNoch keine Bewertungen
- RSA - Brand - Guidelines - 2019 2Dokument79 SeitenRSA - Brand - Guidelines - 2019 2Gigi's DelightNoch keine Bewertungen
- Blue Prism Data Sheet - Provisioning A Blue Prism Database ServerDokument5 SeitenBlue Prism Data Sheet - Provisioning A Blue Prism Database Serverreddy_vemula_praveenNoch keine Bewertungen
- Opc PPT FinalDokument22 SeitenOpc PPT FinalnischalaNoch keine Bewertungen
- WL-80 FTCDokument5 SeitenWL-80 FTCMr.Thawatchai hansuwanNoch keine Bewertungen
- WEB DESIGN WITH AUSTINE-converted-1Dokument9 SeitenWEB DESIGN WITH AUSTINE-converted-1JayjayNoch keine Bewertungen