Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- World IcelandDokument36 SeitenWorld IcelandFranco SantinNoch keine Bewertungen
- Plus2: User ManualDokument64 SeitenPlus2: User ManualFranco SantinNoch keine Bewertungen
- Arabic OCR Arabic OCR Arabic OCR Arabic OCR: User ManualDokument23 SeitenArabic OCR Arabic OCR Arabic OCR Arabic OCR: User ManualFranco SantinNoch keine Bewertungen
- DFF 3Dokument86 SeitenDFF 3Franco SantinNoch keine Bewertungen
- User Manual: TO THEDokument37 SeitenUser Manual: TO THEFranco SantinNoch keine Bewertungen
- Arabic OCR Arabic OCR Arabic OCR Arabic OCR: User ManualDokument23 SeitenArabic OCR Arabic OCR Arabic OCR Arabic OCR: User ManualFranco SantinNoch keine Bewertungen
- Maternal Serum Markers Levels Consecutive Pregnancies:: Possible Genetic Predisposition T o Abnormal LevelsDokument4 SeitenMaternal Serum Markers Levels Consecutive Pregnancies:: Possible Genetic Predisposition T o Abnormal LevelsFranco SantinNoch keine Bewertungen
- Get Unlimited Downloads With A Free Scribd Trial!Dokument2 SeitenGet Unlimited Downloads With A Free Scribd Trial!Franco SantinNoch keine Bewertungen
- Upload A Document - ScribdDokument1 SeiteUpload A Document - ScribdFranco SantinNoch keine Bewertungen
- Publish To The World: Educacion en ArgentinaDokument3 SeitenPublish To The World: Educacion en ArgentinaFranco SantinNoch keine Bewertungen
- Publish To The World: Historia de La Educación Argentina y LatinoamericanaDokument3 SeitenPublish To The World: Historia de La Educación Argentina y LatinoamericanaFranco SantinNoch keine Bewertungen
- Phage L-New Insights Into Regulatory Circuits: Grzegorz We Grzyn, Katarzyna Licznerska, and Alicja We GrzynDokument24 SeitenPhage L-New Insights Into Regulatory Circuits: Grzegorz We Grzyn, Katarzyna Licznerska, and Alicja We GrzynFranco SantinNoch keine Bewertungen
- Publish To The World: Ozlak, Oscar - La Formación Del Estado ArgentinoDokument3 SeitenPublish To The World: Ozlak, Oscar - La Formación Del Estado ArgentinoFranco SantinNoch keine Bewertungen
- Etapas Históricas de La Educación Argentina - RAMALLO - PDF: SearchDokument2 SeitenEtapas Históricas de La Educación Argentina - RAMALLO - PDF: SearchFranco SantinNoch keine Bewertungen
- Publish To The World: Spoken Hindi BookDokument3 SeitenPublish To The World: Spoken Hindi BookReddyNoch keine Bewertungen
- Customs InvoiceDokument1 SeiteCustoms InvoiceFranco Santin100% (1)
- Wiley American Journal of Botany: This Content Downloaded From 181.231.29.59 On Tue, 26 Mar 2019 21:10:07 UTCDokument17 SeitenWiley American Journal of Botany: This Content Downloaded From 181.231.29.59 On Tue, 26 Mar 2019 21:10:07 UTCFranco SantinNoch keine Bewertungen
- The Use of ISSR Markers To Identify Texas Bluegrass Interspecific HybridsDokument3 SeitenThe Use of ISSR Markers To Identify Texas Bluegrass Interspecific HybridsFranco SantinNoch keine Bewertungen
- The Use of ISSR Markers To Identify Texas Bluegrass Interspecific HybridsDokument3 SeitenThe Use of ISSR Markers To Identify Texas Bluegrass Interspecific HybridsFranco SantinNoch keine Bewertungen
- Sex-Linked Markers in Dioecious PlantsDokument7 SeitenSex-Linked Markers in Dioecious PlantsFranco SantinNoch keine Bewertungen
- Choose A Plan, Step 2 of 3 ScribdDokument2 SeitenChoose A Plan, Step 2 of 3 ScribdFranco SantinNoch keine Bewertungen
- Publish To The World: Historia Social y Política de La Educación Argentina TPDokument3 SeitenPublish To The World: Historia Social y Política de La Educación Argentina TPFranco SantinNoch keine Bewertungen
- Bayesian Evolutionary Analysis With BEASTDokument262 SeitenBayesian Evolutionary Analysis With BEASTFranco Santin100% (1)
- Paper Ltketjl Ecje FreioDokument12 SeitenPaper Ltketjl Ecje FreioFranco SantinNoch keine Bewertungen
- Warren 2013Dokument3 SeitenWarren 2013Franco SantinNoch keine Bewertungen
- Wang 2007Dokument6 SeitenWang 2007Franco SantinNoch keine Bewertungen
- Vohra D Sky 2017Dokument21 SeitenVohra D Sky 2017Franco SantinNoch keine Bewertungen
- Paris I 1988Dokument3 SeitenParis I 1988Franco SantinNoch keine Bewertungen
- Shibata 1979Dokument2 SeitenShibata 1979Franco SantinNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- 1 - UK Lamuno Plus BrochureDokument2 Seiten1 - UK Lamuno Plus BrochurePhyo WaiNoch keine Bewertungen
- Kokko - Modelling PDFDokument242 SeitenKokko - Modelling PDFDavid RăscuţoiNoch keine Bewertungen
- Travel With An Ease While Not Putting Uneven Stress On Your Back - Caring Hands PhysiotherapyDokument1 SeiteTravel With An Ease While Not Putting Uneven Stress On Your Back - Caring Hands Physiotherapycaringhands physiotherapyNoch keine Bewertungen
- Group Two'S Seminar Work: Topic: Enzyme Regulation Allosteric Regulation and Models OutlineDokument13 SeitenGroup Two'S Seminar Work: Topic: Enzyme Regulation Allosteric Regulation and Models OutlineOluwasegun ModupeNoch keine Bewertungen
- P.E & Health 11Dokument8 SeitenP.E & Health 11Yvonnie TamayoNoch keine Bewertungen
- Dna Report NEWDokument21 SeitenDna Report NEWshivankdwivedi3Noch keine Bewertungen
- Ospe-Osce Microbio-Pathology SlidesDokument208 SeitenOspe-Osce Microbio-Pathology Slidesdoterofthemosthigh100% (3)
- Kombinasi Pengaruh Media Tanam Akar Pakis Dan Arang Sekam Terhadap Perkecambahan Dan Pertumbuhan Bibit EucalyptusDokument9 SeitenKombinasi Pengaruh Media Tanam Akar Pakis Dan Arang Sekam Terhadap Perkecambahan Dan Pertumbuhan Bibit EucalyptusNeta AdzkiyaNoch keine Bewertungen
- Cambridge International AS & A Level: BIOLOGY 9700/35Dokument16 SeitenCambridge International AS & A Level: BIOLOGY 9700/35photopour13sNoch keine Bewertungen
- ANIMILIADokument10 SeitenANIMILIAAnand Teekmani AnuNoch keine Bewertungen
- EXPERIMENT 9 Food ChemistryDokument7 SeitenEXPERIMENT 9 Food ChemistryNurmazillazainal67% (3)
- Lesson-Plan Ecologica RelationshipDokument10 SeitenLesson-Plan Ecologica RelationshipRen Mart Decipeda100% (3)
- Toxicity of Clay Occulation of The Toxic Dino Agellate, Karenia Brevis, To Estuarine Invertebrates and SHDokument12 SeitenToxicity of Clay Occulation of The Toxic Dino Agellate, Karenia Brevis, To Estuarine Invertebrates and SHinousseNoch keine Bewertungen
- Of Basques, Blood, and Blue PeopleDokument6 SeitenOf Basques, Blood, and Blue PeopleVince MiglioreNoch keine Bewertungen
- La Teoría de La Compuerta - AlDokument12 SeitenLa Teoría de La Compuerta - AlAlterOsPegiamNoch keine Bewertungen
- Skeletal Muscle Relaxant (Lecture Notes QIUP 2017)Dokument9 SeitenSkeletal Muscle Relaxant (Lecture Notes QIUP 2017)Monissha ThiaguNoch keine Bewertungen
- Bio 14 NotesDokument5 SeitenBio 14 Noteshey lungNoch keine Bewertungen
- ProteinsDokument43 SeitenProteinsBilalNoch keine Bewertungen
- Paper 16 Vinay Malik PDFDokument5 SeitenPaper 16 Vinay Malik PDFPRABHAT KUMAR CHAUHANNoch keine Bewertungen
- 266 1531 2 PBDokument8 Seiten266 1531 2 PBMariatul kiftia UsemahuNoch keine Bewertungen
- Goby Shirmp MutualismDokument20 SeitenGoby Shirmp MutualismKhandoker Raisul AzadNoch keine Bewertungen
- Tissue Engineered Scaffold Fabrication Methods For Medical ApplicationsDokument26 SeitenTissue Engineered Scaffold Fabrication Methods For Medical ApplicationsSarah FathiNoch keine Bewertungen
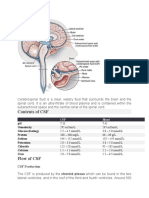
- Cerebrospinal FluidDokument3 SeitenCerebrospinal FluidlecturioNoch keine Bewertungen
- Review of LaboratoryDokument48 SeitenReview of LaboratoryIkhar RidhoNoch keine Bewertungen
- Esthetic Dentistry / Orthodontic Courses by Indian Dental AcademyDokument256 SeitenEsthetic Dentistry / Orthodontic Courses by Indian Dental Academyindian dental academyNoch keine Bewertungen
- Research Paper On 3D Printed Prosthesis As A Better AlternativeDokument16 SeitenResearch Paper On 3D Printed Prosthesis As A Better AlternativeTanbin Rafee100% (1)
- Corrections and Additions To Taxonomists' Glossary of Mosquito AnatomyDokument17 SeitenCorrections and Additions To Taxonomists' Glossary of Mosquito AnatomymokarahNoch keine Bewertungen
- 5991 7951en PDFDokument12 Seiten5991 7951en PDFAlonso Hurtado100% (1)
- The Tale of MR Jeremy FisherDokument95 SeitenThe Tale of MR Jeremy FisherTrent DouglasNoch keine Bewertungen
- The Flavor and Fragrance High Production Volume ConsortiaDokument33 SeitenThe Flavor and Fragrance High Production Volume Consortiaesteban4567Noch keine Bewertungen