Das könnte Ihnen auch gefallen
- Phylogeny and Evolution of the AngiospermsVon EverandPhylogeny and Evolution of the AngiospermsBewertung: 5 von 5 Sternen5/5 (1)
- Evolution: An Overview Based on Genetic Characters and BirdsVon EverandEvolution: An Overview Based on Genetic Characters and BirdsNoch keine Bewertungen
- Great Moments in Evolthe Conquest of LandDokument6 SeitenGreat Moments in Evolthe Conquest of LandClaraNoch keine Bewertungen
- PCW 023Dokument5 SeitenPCW 023Federico GarciaNoch keine Bewertungen
- Marine Ecology - 2011 - Cecere - Vegetative Reproduction by Multicellular Propagules in Rhodophyta An OverviewDokument19 SeitenMarine Ecology - 2011 - Cecere - Vegetative Reproduction by Multicellular Propagules in Rhodophyta An OverviewKathya Gómez AmigoNoch keine Bewertungen
- Kenrick 2017-Ciclos de Vida en Rhynia Chert PDFDokument12 SeitenKenrick 2017-Ciclos de Vida en Rhynia Chert PDFMia ScNoch keine Bewertungen
- American J of Botany - 2004 - Shaw - Phylogeny and Diversification of BryophytesDokument25 SeitenAmerican J of Botany - 2004 - Shaw - Phylogeny and Diversification of BryophytesVijayakumarNoch keine Bewertungen
- Plant BIology - Plant EvolutionDokument6 SeitenPlant BIology - Plant EvolutionRhain GarciaNoch keine Bewertungen
- Phylogeny and Diversification of BryophytesDokument26 SeitenPhylogeny and Diversification of BryophytesHamengku AgengNoch keine Bewertungen
- Ageing Research Reviews: Wouter G. Van Doorn, Kohki YoshimotoDokument14 SeitenAgeing Research Reviews: Wouter G. Van Doorn, Kohki YoshimotovinoymaniNoch keine Bewertungen
- The First Land Plants: EVOLUTIONARY BIOLOGY: A Plant PerspectiveDokument9 SeitenThe First Land Plants: EVOLUTIONARY BIOLOGY: A Plant PerspectiveNindzRMapeNoch keine Bewertungen
- Algal Phylogeny and The Origin of Land Plants: Update On EvolutionDokument7 SeitenAlgal Phylogeny and The Origin of Land Plants: Update On EvolutionGaston PerezNoch keine Bewertungen
- De Vries - Archibald - 2018 - Plant Evolution - Landmarks On The Path To Terrestrial LifeDokument7 SeitenDe Vries - Archibald - 2018 - Plant Evolution - Landmarks On The Path To Terrestrial LifeFernando AndradeNoch keine Bewertungen
- Desarrollo Del CloroplastoDokument21 SeitenDesarrollo Del CloroplastoAnderson HernándezNoch keine Bewertungen
- Fpls 12 735020Dokument15 SeitenFpls 12 735020Maria Tereza Faria FariaNoch keine Bewertungen
- Shaw, J. & K. Renzaglia. (2004) - Phylogeny and Diversification of Bryophytes. American PDFDokument26 SeitenShaw, J. & K. Renzaglia. (2004) - Phylogeny and Diversification of Bryophytes. American PDFDiego Andres Moreno GaonaNoch keine Bewertungen
- Ecological and Economic Significance of BryophytesDokument26 SeitenEcological and Economic Significance of BryophytesbukundadaNoch keine Bewertungen
- Effects of The Harmful Dinoflagellate Ostreopsis Cf. Ovata On Different Life Cycle Stages of The Common Moon Jellyfish Aurelia Sp.Dokument10 SeitenEffects of The Harmful Dinoflagellate Ostreopsis Cf. Ovata On Different Life Cycle Stages of The Common Moon Jellyfish Aurelia Sp.Cintya NevarezNoch keine Bewertungen
- Phylogenomic Evidence For The Monophyly of Bryophytes and The Reductive Evolution of StomataDokument15 SeitenPhylogenomic Evidence For The Monophyly of Bryophytes and The Reductive Evolution of StomataROBERTO MIRANDA CALDASNoch keine Bewertungen
- PerkecambahanDokument21 SeitenPerkecambahanTT ZmDESNoch keine Bewertungen
- Journal of Phycology - 2003 - Grzebyk - THE MESOZOIC RADIATION OF EUKARYOTIC ALGAE THE PORTABLE PLASTID HYPOTHESIS1Dokument9 SeitenJournal of Phycology - 2003 - Grzebyk - THE MESOZOIC RADIATION OF EUKARYOTIC ALGAE THE PORTABLE PLASTID HYPOTHESIS1obaytutNoch keine Bewertungen
- Ecological and Economic Significance of BryophytesDokument26 SeitenEcological and Economic Significance of BryophytesMohammad ShakibNoch keine Bewertungen
- Greenwood 1991. The Taphonomy of Plant MacrofossilsDokument29 SeitenGreenwood 1991. The Taphonomy of Plant MacrofossilsDiana ArroyoNoch keine Bewertungen
- Bryophytes: The Leap Onto LandDokument27 SeitenBryophytes: The Leap Onto LandaniruskNoch keine Bewertungen
- Molecular Phylogenetics and EvolutionDokument14 SeitenMolecular Phylogenetics and EvolutionNadin AriaNoch keine Bewertungen
- Delavault - 2015 - Knowing The Parasite Biology and Genetics of OrobDokument16 SeitenDelavault - 2015 - Knowing The Parasite Biology and Genetics of OrobJustin PawlonskiNoch keine Bewertungen
- Review: Field Guide To Plant Model SystemsDokument15 SeitenReview: Field Guide To Plant Model SystemsRachnaNoch keine Bewertungen
- Ch29 PlantDiversityI1Dokument14 SeitenCh29 PlantDiversityI1Cristie SumbillaNoch keine Bewertungen
- Casey McGrath 2020Dokument2 SeitenCasey McGrath 2020Ravindi SamarasingheNoch keine Bewertungen
- Agrawal - 2012Dokument21 SeitenAgrawal - 2012EduardoDeOliveiraBastosNoch keine Bewertungen
- Macffaen 2001. Primary and Secondary Endosymbiosis and The OriginDokument10 SeitenMacffaen 2001. Primary and Secondary Endosymbiosis and The OriginGabyNoch keine Bewertungen
- Tribble Et Al. 2019Dokument17 SeitenTribble Et Al. 2019julietanatteroNoch keine Bewertungen
- To Cite This Article: K. Evans & A. R. Stone (1977) A Review of The Distribution and Biology of The Potato CystDokument13 SeitenTo Cite This Article: K. Evans & A. R. Stone (1977) A Review of The Distribution and Biology of The Potato Cystabdneour zaouaguiNoch keine Bewertungen
- 25.1 Early Plant Life: Algae and Evolutionary Paths To PhotosynthesisDokument7 Seiten25.1 Early Plant Life: Algae and Evolutionary Paths To Photosynthesisklavier10244379Noch keine Bewertungen
- Kingdom Plantae Tree of LifeDokument3 SeitenKingdom Plantae Tree of LifekathluqNoch keine Bewertungen
- Nature 08057Dokument8 SeitenNature 08057Raghav PathakNoch keine Bewertungen
- Bradyrhizobium e o Que Nodula, Varias ReferenciasDokument17 SeitenBradyrhizobium e o Que Nodula, Varias ReferenciasJordanaNoch keine Bewertungen
- 1 s2.0 S0960982218309072 MainDokument18 Seiten1 s2.0 S0960982218309072 MainYi-Ling ChiuNoch keine Bewertungen
- Los Origenes de Las Plantas Con Flores y PolinizadoresDokument4 SeitenLos Origenes de Las Plantas Con Flores y PolinizadoresRuve20Noch keine Bewertungen
- Substratum Preference of Two Species of Xanthoparmelia: Chris DEDUKE, Michele D. Piercey-NormoreDokument11 SeitenSubstratum Preference of Two Species of Xanthoparmelia: Chris DEDUKE, Michele D. Piercey-NormoreGraciela Anabel Castro ChumbeNoch keine Bewertungen
- Kelling 2009 CromoalveoladosDokument8 SeitenKelling 2009 CromoalveoladosGabyNoch keine Bewertungen
- Chapter 25. Seedless PlantsDokument76 SeitenChapter 25. Seedless PlantsRicardo RicoNoch keine Bewertungen
- Biosynthetic Origin of Natural Products Isolated From Marine Microorganism-Invertebrate AssemblagesDokument8 SeitenBiosynthetic Origin of Natural Products Isolated From Marine Microorganism-Invertebrate Assemblagesashok1anand2Noch keine Bewertungen
- Green Algae and The Origin of Land PlantsDokument22 SeitenGreen Algae and The Origin of Land PlantssaporettiNoch keine Bewertungen
- 4 AfcafDokument16 Seiten4 AfcafYi-Ling ChiuNoch keine Bewertungen
- Temporal Variation of Dinoflagellate Species Composition Along Fish Cage Farm at Santubong RiverDokument52 SeitenTemporal Variation of Dinoflagellate Species Composition Along Fish Cage Farm at Santubong RiverNadia RajamiNoch keine Bewertungen
- Embryophyte - WikipediaDokument73 SeitenEmbryophyte - WikipediaBashiir NuurNoch keine Bewertungen
- Mcelwain 2007Dokument10 SeitenMcelwain 2007Trần Minh HuyNoch keine Bewertungen
- Symbiodinium Necroappetens Sp. Nov. Dinophyceae An Opportunist Zooxanthella Found in Bleached and Diseased Tissues of Caribbean Reef CoralsDokument17 SeitenSymbiodinium Necroappetens Sp. Nov. Dinophyceae An Opportunist Zooxanthella Found in Bleached and Diseased Tissues of Caribbean Reef CoralsThu Anh NguyenNoch keine Bewertungen
- Chloroplast SymbiosisDokument10 SeitenChloroplast SymbiosisMaria Quesada BrunaNoch keine Bewertungen
- Plant Roots: Understanding Structure and Function in An Ocean of ComplexityDokument6 SeitenPlant Roots: Understanding Structure and Function in An Ocean of ComplexityEthan BalakrishnanNoch keine Bewertungen
- Brun Et Al. - 2018 - Seed Germination in Parasitic Plants What InsightDokument16 SeitenBrun Et Al. - 2018 - Seed Germination in Parasitic Plants What InsightJustin PawlonskiNoch keine Bewertungen
- Implications For Arthropod Fossil RecordDokument9 SeitenImplications For Arthropod Fossil RecordRamon Estadella SerraNoch keine Bewertungen
- Freshwater Biology - 2023 - GreenDokument18 SeitenFreshwater Biology - 2023 - GreenTITO FAUZAN ADHIMNoch keine Bewertungen
- Early Land Plants 2019Dokument17 SeitenEarly Land Plants 2019Joseane FerrazNoch keine Bewertungen
- The Trials and Tribulations of The Plant Male Gametophyte - Understanding Reproductive Stage Stress ToleranceDokument54 SeitenThe Trials and Tribulations of The Plant Male Gametophyte - Understanding Reproductive Stage Stress ToleranceArya ChowdhuryNoch keine Bewertungen
- The Trials and Tribulations of The Plant Male Gametophyte - Understanding Reproductive Stage Stress ToleranceDokument54 SeitenThe Trials and Tribulations of The Plant Male Gametophyte - Understanding Reproductive Stage Stress ToleranceArya ChowdhuryNoch keine Bewertungen
- The Nature of Macroalgae and Their Interactions On Reefs: Mark M. Littler and Diane S. LittlerDokument12 SeitenThe Nature of Macroalgae and Their Interactions On Reefs: Mark M. Littler and Diane S. LittlerFrankie AndrewsNoch keine Bewertungen
- Symbiotic InteractionsDokument18 SeitenSymbiotic InteractionsJotha-OriginNoch keine Bewertungen
- Plaant WorldDokument10 SeitenPlaant WorldBallen ZayrNoch keine Bewertungen
- Duavent Drug Study - CunadoDokument3 SeitenDuavent Drug Study - CunadoLexa Moreene Cu�adoNoch keine Bewertungen
- SHCDokument81 SeitenSHCEng Mostafa ElsayedNoch keine Bewertungen
- Technical Specification For 33KV VCB BoardDokument7 SeitenTechnical Specification For 33KV VCB BoardDipankar ChatterjeeNoch keine Bewertungen
- Emea 119948060Dokument31 SeitenEmea 119948060ASHUTOSH MISHRANoch keine Bewertungen
- FM 2030Dokument18 SeitenFM 2030renaissancesamNoch keine Bewertungen
- Lecture 4 ENGR 243 DynamicsDokument45 SeitenLecture 4 ENGR 243 DynamicsRobby RebolledoNoch keine Bewertungen
- An Exploration of The Ethno-Medicinal Practices Among Traditional Healers in Southwest Cebu, PhilippinesDokument7 SeitenAn Exploration of The Ethno-Medicinal Practices Among Traditional Healers in Southwest Cebu, PhilippinesleecubongNoch keine Bewertungen
- Arnold Ventures Letter To Congressional Social Determinants of Health CaucusDokument7 SeitenArnold Ventures Letter To Congressional Social Determinants of Health CaucusArnold VenturesNoch keine Bewertungen
- Monergism Vs SynsergismDokument11 SeitenMonergism Vs SynsergismPam AgtotoNoch keine Bewertungen
- Negotiating Skills Negotiating Skills: To Provide You With The Skills To Plan & Implement Successful NegotiationDokument32 SeitenNegotiating Skills Negotiating Skills: To Provide You With The Skills To Plan & Implement Successful NegotiationKanimozhi.SNoch keine Bewertungen
- The Magic DrumDokument185 SeitenThe Magic Drumtanishgiri2012Noch keine Bewertungen
- From Philo To Plotinus AftermanDokument21 SeitenFrom Philo To Plotinus AftermanRaphael888Noch keine Bewertungen
- My Personal Code of Ethics1Dokument1 SeiteMy Personal Code of Ethics1Princess Angel LucanasNoch keine Bewertungen
- 1.technical Specifications (Piling)Dokument15 Seiten1.technical Specifications (Piling)Kunal Panchal100% (2)
- Service Manual: SV01-NHX40AX03-01E NHX4000 MSX-853 Axis Adjustment Procedure of Z-Axis Zero Return PositionDokument5 SeitenService Manual: SV01-NHX40AX03-01E NHX4000 MSX-853 Axis Adjustment Procedure of Z-Axis Zero Return Positionmahdi elmay100% (3)
- Hamstring - WikipediaDokument21 SeitenHamstring - WikipediaOmar MarwanNoch keine Bewertungen
- Tribal Banditry in Ottoman Ayntab (1690-1730)Dokument191 SeitenTribal Banditry in Ottoman Ayntab (1690-1730)Mahir DemirNoch keine Bewertungen
- MASONRYDokument8 SeitenMASONRYJowelyn MaderalNoch keine Bewertungen
- UTP Student Industrial ReportDokument50 SeitenUTP Student Industrial ReportAnwar HalimNoch keine Bewertungen
- All You Need To Know About Egg YolkDokument7 SeitenAll You Need To Know About Egg YolkGolden Era BookwormNoch keine Bewertungen
- PR KehumasanDokument14 SeitenPR KehumasanImamNoch keine Bewertungen
- Project - New Restuarant Management System The Grill HouseDokument24 SeitenProject - New Restuarant Management System The Grill HouseMayank Mahajan100% (3)
- Chapter13 PDFDokument34 SeitenChapter13 PDFAnastasia BulavinovNoch keine Bewertungen
- Chapter 1 ClassnotesDokument35 SeitenChapter 1 ClassnotesAllison CasoNoch keine Bewertungen
- Traveling Salesman ProblemDokument11 SeitenTraveling Salesman ProblemdeardestinyNoch keine Bewertungen
- V737 OverheadDokument50 SeitenV737 OverheadnewahNoch keine Bewertungen
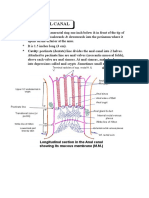
- Anatomy Anal CanalDokument14 SeitenAnatomy Anal CanalBela Ronaldoe100% (1)
- PLC Laboratory Activity 2Dokument3 SeitenPLC Laboratory Activity 2Kate AlindajaoNoch keine Bewertungen
- Talking Art As The Spirit Moves UsDokument7 SeitenTalking Art As The Spirit Moves UsUCLA_SPARCNoch keine Bewertungen
- DC 7 BrochureDokument4 SeitenDC 7 Brochures_a_r_r_yNoch keine Bewertungen